Abstract
The COVID-19 has started from China and spread to all countries within a very short period. The severity of the disease varies from one patient to another as well as one country to another. It depends on the immune status of individuals simultaneously on the quality of the environment and customs of a particular country. Various factors are studied in correlation with the severity of disease like specific dietary supplements, previous exposure to related antigens, age, and health of individual and previous vaccinations, and many more. Different vaccines play an essential role to reduce the severity of various infectious diseases like TB, Measles, etc. Few bacterial vaccines also showed non-specific activity against viral infections like the BCG vaccine. The efficacy of BCG vaccine against several viral infections reported in several countries like West Africa, South Africa, Australia, etc. BCG was found to be associated with the reduction in children’s mortality rate due to respiratory infections in Haiti, Southeast Asia, United Kingdom, etc. This study focused on BCG vaccination concerning the severity of disease among the vaccinated and non-vaccinated population. Further, the study explained the role of BCG in viral inactivation with emphasis on the COVID-19 virus. We found low severity of SARSCoV- 2 infections in universal BCG implemented countries as compare to the non-universal implemented countries. The current study explored that the BCG vaccine is showing a strong correlation in the reduction of the severity of COVID-19 infection.
Keywords
BCG, COVID-19, Non-specific, Immune response, Trained-immunity, Cytokines
Introduction
Bacille Calmette Guerin (BCG) vaccine has existence of more than 80 years with its widest use among all currently available vaccines [1]. BCG vaccine developed from a virulent attenuated M. bovis strain against tuberculosis (TB) by deleting more than 14 different regions in the genome of Mycobacterium [2]. Several countries are having high prevalence rate of TB, BCG vaccine is used as preventive measure for tubercle meningitis in children. The countries with low risk usually do not include BCG vaccine in their vaccination schedule. BCG is administered immediately to new-borns within a few hours or days of birth. BCG vaccine has 70% potency against meningitis and miliary tuberculosis, whereas the efficacy against pulmonary TB, 52% was reported [3-6]. The intracellular surviving ability of Mycobacteria in phagocytic cells has been tackled with the development of a vaccine with front focus on cell-mediated immunity [7]. The innate immunity being the barrier for every infection has proved to reduce the severity with early recovery. Phagocytosis is the wellestablished system for the recognition and killing of microorganisms [7]. Various studies have shown the role of BCG vaccine in multiple signalling pathways that have the potent to stimulate the innate myeloid and lymphoid cellular population. Dendritic cells, macrophages, monocytes and neutrophils are activated by BCG vaccine and eventually induce T cell mediated immunity [8,9]. Several studies have shown the reduction in hospitalization due to respiratory infections in case of BCG vaccinated individuals. BCG vaccination has been reduced child mortality all over the world, including Haiti, India, Sweden, South or Southeast Asia, and the United Kingdom [10-13]. A high childhood mortality rate in Guinea-Bissau associated with unclassified fever or malaria associated to be diminished with BCG-vaccination.
BCG Non-specific Immune Response
BCG vaccine stimulates the innate immune response or cell-mediated immune response against similar types of pathogens by developing the immune memory. Several studies proved that the BCG vaccine protects from various types of infections like parasitic, bacterial, and viral by its non-specific activity for the pathogens causing pneumonialike pathology. Maternal vaccination against the measles might be associated with child mortality reduction by maternal antibodies. This might be sharing the same mechanism, which acts in case of BCG to enhance the nonspecific efficacy [14]. Intravenous injection of BCG in mice has reported to protect against Babesia and Plasmodium species and proved that it is not associated with specific antibody [15,16]. BCG may have role in the protection beyond the target pathogens that could elevate through the memory of innate immune response or heterologous lymphocyte activation. BCG vaccine has also shown a role in the reduction of acute lower respiratory tract infections (ALRI) risk among vaccinated children as compared to the non-vaccinated child population [17,18]. Apart from TB, the BCG vaccine also showed a specific role in children’s fatality rate reduction by its non-specific potential against unrelated pathogens [10,19]. Effect of the BCG vaccine in other respiratory tract infections reported in the older Indonesian population [20]. A clinical trial conducted in Japan on the BCG vaccination in aged tuberculinnegative population with pneumonia that shows adequate protection against pneumonia [21]. Several studies showed a 40% reduction in mortality by malaria, sepsis, respiratory infections, and leprosy in BCG vaccinated people, conducted in West Africa [22-26]. A study conducted in the South African adolescent population also showed that the BCG vaccine reduces upto 70% respiratory tract infections [27].
A study reported that BCG vaccinated infants’ exhibit upregulated IgG production against the antigens of the hepatitis B vaccine [28]. BCG vaccination showed the association with IgG’s high titer against H-influenzae type b polysaccharide, pneumococcal capsular polysaccharide antigens, and tetanus toxoid in vaccinated and non- BCG vaccinated Australian’s infants’ population [29]. Clinical trials also showed that BCG decreases unrelated respiratory tract infections [21,30]. Moreover, viruses are specific for their hosts with intracellular obligate parasitic nature. Viruses become more pathogenic when these change the host or undergo antigenic mutations. Innate immune response might eliminate the viruses and virusinfected cells. Various clinical trials launched intending to generate data from all around the world to find out the most efficient treatment for COVID-19. However, it is not possible to prevent the spread of any infection through the vaccination. Vaccination might be able to increase the immune response by reducing the severity of infections. SARS viruses also reported in the respiratory tract related medical conditions like inflammation, pneumonia, etc., and BCG already reported in the reduction of respiratory tract related infection after vaccination. A study reported that BCG has the potential to protect individuals from infection up to 60 years of age. A high mortality rate has found among the aged and co-morbid population infected with SARS-CoV-2 [31]. The mechanism of innate immune response plays an essential role for the elimination of pathogens. The versatile existence and diversity at every level of life in evolutionary tree has set up its well-marked role.
BCG Induced Immune Response
BCG involves in the stimulation of Pathogen Recognition Receptors (PRRs) like Toll-like receptors (TLR) receptors and nucleotide-binding oligomer domain-like receptors (NOD) [8,32] and activates the innate immune system. BCG antigens mainly act on TLR2, TLR4, TLR7/8 and TLR9 receptors and initiate the activation of macrophages and DCs later on maturation. TLRs further start the cascade through the activation of the MYD88 adapter molecule, which helps in the TRAF6 (TNF receptor-associated factor 6), IRAK (IL1 receptor-associated kinase), and MAPK (Mitogen-activated protein kinase) activations. TRAF6, IRAK, and MAPK consequently activate the nuclear transcription factor (NF-kB), which activates the genes associated with the secretion of the pro-inflammatory cytokine [33].
The mechanism of BCG vaccine-induced immune response and stimulates the c chemokine and cytokines secretion in the microenvironment that besides spurs other immune cells in a non-specific manner. BCG vaccine stimulates the secretion of different cytokines like IL-2, TNF-α, and IFN-γ by the activation of macrophages and T cells. IFN-γ secreting T cells showed the CCR7+ CD62L phenotype expressing the central memory T cells in an invitro experiment [34] and also found in neonates till 27 weeks of BCG vaccination [35]. However, BCG vaccination also stimulates the smaller subset of T effector cells. IFN-γ secretion regulates by the IL-10 secreted through T regulatory cells (Tregs). BCG might activate the Tregs with CD4+CD25+ phenotype that acts non-specifically against non-TB related health problems [36]. Tregs cells stimulated by BCG secrete cytokines such as TGF-β and IL-10 [37]. Sebina et. al., have shown that the PPD-specific IFN-γ levels are higher in BCG vaccinated individuals whereas lower in unvaccinated individuals. The study has proved the robust activation of long-lived T cells in association with BCG vaccination [38]. Professional Antigen Presenting Cells (APCs) represent BCG antigens to the helper T-cells by using the MHC molecules. These interactions need to expressions of both types of MHCs molecules identified through TCR of CD4+ T-cells and CD8+ T-cells [39]. T-cells differentiation into Th-17 and Th-22 depends on the levels of cytokines and their downstream signaling. Th-17 cells are differentiated from T-cells by the IL-6, IL-23, and TGFβ cytokines actions, which produce the cytokine IL-17. Whereas, Th-22 cells differentiate by the action of IL-6 and TNF-α on Th-cells that secrets IL- 22 [40]. BCG vaccine also stimulates the differentiation of mononuclear cells into a specific group of cells that are BCG Activated Killer cells (BAK) [41]. Targeted killing cells are lymphocyte-activated killer cells (LAK), CD8+, and CD56+ that help in the clearance of pathogens. Th-1 cytokines such as IL-2 and IFN-γ activate the BAK cells for cytotoxicity [41-43] (Figure 1 [44]).
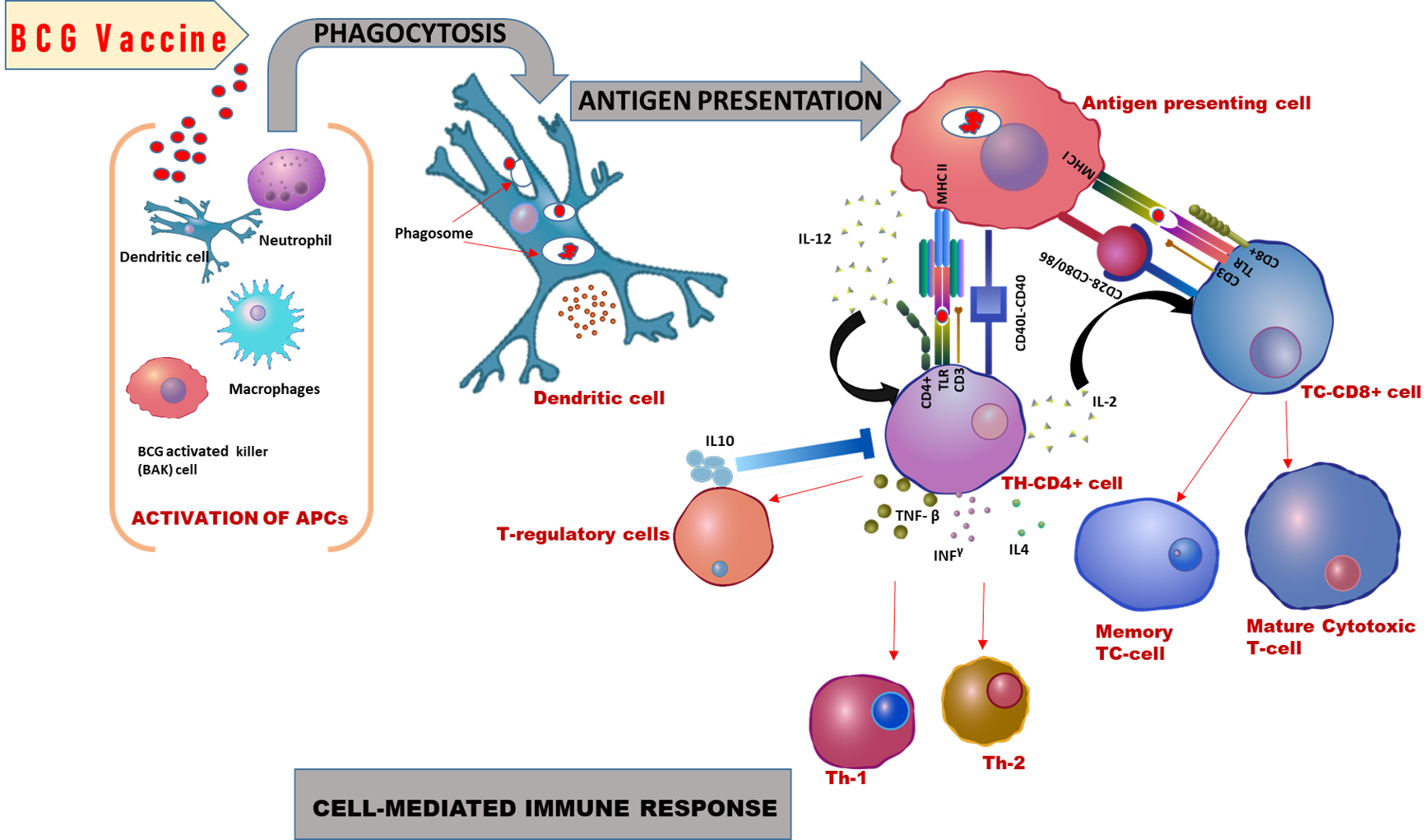
BCG vaccine helps in the reduction of inflammation and symptoms of infectious diseases by inhibiting the inflammasome activation via Zinc-metalloproteases [45]. Zinc signaling exhibits an essential role in the immune response against viral infections by maintaining the proper immune functions [46]. A study showed that the BCG mutant zmp1 deletion enhanced immunogenicity by increased IFN-γ production, antigen presentation, and DTH [47]. Currently, physicians prescribing the Zinc consumption in coronavirus infection that might help to reduce the disease pathology and severity of the infection. Moreover, the BCG vaccine already activated the similar immunogenicity in the vaccinated population.
Immune Activation against SARS-CoV-2
The PRRs of the TLR family or DExD/H box RNA helicase family initiates the immune response inside the host upon virus entry. The cell receptors on activation give a signal to increase the levels of IFNs, especially IFN-α, and INF-β, and various other cytokines [48]. Different TLRs are inducible although, some are constitutively present on specialized immune cells like dendritic cells for the rapid production of IFNs and cytokines [49].
The TLRs molecules are too selective for their purpose, like to recognition of specific viral particles. TLR7/8 receptors bind with the single ssRNA and initiate the immune response with inflammatory cytokines secretion. TLR3 receptors are specific for double-stranded RNA (dsRNA) as a ligand and form an active signaling complex that initiates the immune response [50-54]. SARS viruses activate the immune system in the same way as the BCG vaccine induces. SARS-CoV-2 activates the immune system through the TLRs and PAMPs [55]. The innate and adaptive immune responses occur against the SARS infections through the phagocytic process of macrophages and antigen presentation to the naive T-cells. It starts T-cells activation and proliferation with differentiation by the secretion of various types of cytokines that help in the differentiation of T-cells sub-groups like Th-17, etc. The huge cytokines production increases the amplification of immune response. The constant production of these mediators due to the viral persistency might alter the CD8+ T cell Natural-Killer cell activation. Though, CD8+ T-cells are the most powerful mediators in cell-mediated immune actions to the clearance of SARS-CoV-2. Viral dsRNA might initiate the immune response during CoV replication via TLR3 signaling cascades like IRFs and NF-κB activation inside the infected cells or antigenpresenting cells. These cells produce a large amount of type I IFNs and pro-inflammatory cytokines [56,57]. For the protection of uninfected cells from the viral proteins, type-I IFNs plays a significant role in the clearance of the viral proteins. Essential proteins of SARS viruses are interfered with in TLRs signaling and try to escape the immune response via TLRs activation [54,58]. Viral spike proteins are recognized by the TLRs and get activated that stimulate pro-inflammatory cytokines secretion by using MyD88-dependent pathways. Virus and host-cell interactions enhance the immune mediators’ secretions like chemokines and cytokines (IL-1, IL-2, etc.). Chemokines and cytokines help in activation of leukocytes and lymphocytes. Thus, activating the immune system against the SARS-CoV-2 infection. BCG vaccine already initiated that type of immune response in the BCG vaccinated population, which might be activated by the SARS viruses. Hence BCG vaccine decreases the time of immune response against the COVID-19 and reducing the severity of the infection with an increasing rate of recovery [59] (Figure 2 [44]).
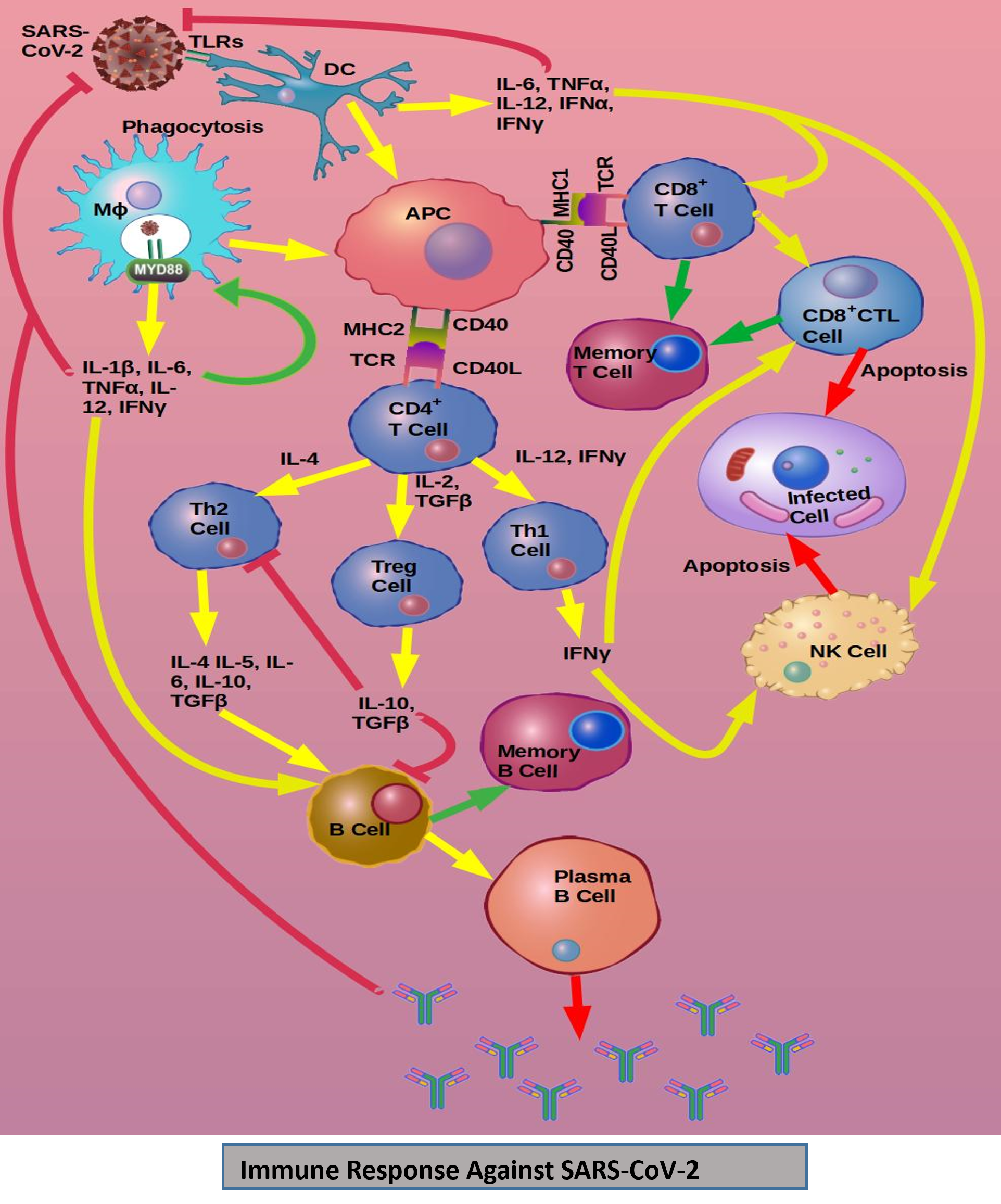
The Data of SARS-CoV-2 infections, severity, and recovery has shown a strong association among BCG vaccination. The universal BCG implemented countries are showing the very low severity and high rate of recovery as compared to the not universally BCG vaccination implemented countries. BCG trained immunity against the SARS-CoV-2 might have a role in the mechanism by which induces its beneficial effects. BCG vaccination has shown the reduction of viremia as compared to yellow fever vaccination (live attenuated) in healthy human volunteers. The immune response in healthy human volunteers might be associated with epigenetic changes in monocytes that could help to improve the antiviral responses [60]. Trained immunity of BCG might have a role in important protection against various viral infections. Thus, the BCG vaccine could play an important role in the reduction of SARS-CoV-2 infection severity and recovery in universally BCG implemented countries.
Status of SARS-CoV-2 in Vaccinated and Non-vaccinated Countries
By date 28 August 2020, the status of SARS-CoV-2 positive cases in countries vaccinated with BCG are 16,755,837, mortality in percentage is 3%, and recovery in percentage is 76.256%, whereas, in non-vaccinated countries, the SARS-CoV-2 positive cases are 7,751,688, mortality in percentage is 4.13%, and recovery rate is 52.45%. Thus, the data showed that BCG might play an essential role in the reduction of deaths by COVID-19 and increase recovery [61].
Conclusion
The BCG vaccine is the oldest vaccine that is being used against TB in all over the world. The efficacy of BCG to boost immunity against M. Tuberculosis is not 100 percent, but it also protects from other pathogens like bacteria, fungi, and viruses. It activates the innate immune response and cell-mediated immune response, which is related to the non-specific efficacy of BCG. The nonspecific immune response is significantly essential to clear viral-like pathogens. BCG vaccine dominantly activates the cell-mediated immunity via TLRs and IFNs, and it also develops the memory against similar types of pathogens, which cause TB like symptoms such as pneumonia. Several studies have reported the BCG vaccine’s nonspecific role against various viral infections. Currently, the infection of SARS-CoV-2 is less symptomatic in the population vaccinated with BCG at all over the globe. The SARS-CoV-2 virus severity decreases, whereas recovery increases in vaccinated countries populations as compared to the non-vaccinated.
References
2. Brosch R, Pym AS, Gordon SV, Cole ST. The evolution of mycobacterial pathogenicity: clues from comparative genomics. Trends in Microbiology. 2001 Sep 1;9(9):452-8.
3. Colditz GA, Berkey CS, Mosteller F, Brewer TF, Wilson ME, Burdick E, et al. The efficacy of bacillus Calmette- Guerin vaccination of newborns and infants in the prevention of tuberculosis: meta-analyses of the published literature. Pediatrics. 1995 Jul 1;96(1):29-35.
4. Miceli I, De Kantor IN, Colaiácovo D, Peluffo G, Cutillo I, Gorra R, Botta R, et al. Evaluation of the effectiveness of BCG vaccination using the case control method in Buenos Aires, Argentina. International Journal of Epidemiology. 1988 Sep 1;17(3):629-34.
5. Trunz BB, Fine PE, Dye C. Effect of BCG vaccination on childhood tuberculous meningitis and miliary tuberculosis worldwide: a meta-analysis and assessment of costeffectiveness. The Lancet. 2006 Apr 8;367(9517):1173-80.
6. Bonifachich E, Chort M, Astigarraga A, Diaz N, Brunet B, Pezzotto SM, et al. Protective effect of bacillus Calmette-Guerin (BCG) vaccination in children with extrapulmonary tuberculosis, but not the pulmonary disease: a case-control study in Rosario, Argentina. Vaccine. 2006 Apr 5;24(15):2894-9.
7. Trunz BB, Fine PE, Dye C. Effect of BCG vaccination on childhood tuberculous meningitis and miliary tuberculosis worldwide: a meta-analysis and assessment of costeffectiveness. The Lancet. 2006 Apr 8;367(9517):1173-80.
8. Jasenosky LD, Scriba TJ, Hanekom WA, Goldfeld AE. T cells and adaptive immunity to Mycobacterium tuberculosis in humans. Immunological Reviews. 2015 Mar;264(1):74-87.
9. Bickett TE, McLean J, Creissen E, Izzo L, Hagan C, Izzo AJ, et al. Characterizing the BCG Induced Macrophage and Neutrophil Mechanisms for Defense Against Mycobacterium tuberculosis. Frontiers in Immunology. 2020 Jun 18;11:1202.
10. Kumar P, Tyagi R, Das G, Bhaskar S. Mycobacterium indicus pranii and Mycobacterium bovis BCG lead to differential macrophage activation in T oll-like receptordependent manner. Immunology. 2014 Oct;143(2):258- 68.
11. de Castro MJ, Pardo-Seco J, Martinón-Torres F. Nonspecific (heterologous) protection of neonatal BCG vaccination against hospitalization due to respiratory infection and sepsis. Clinical Infectious Diseases. 2015 Jun 1;60(11):1611-9.
12. Shann F. The non-specific effects of vaccines. Archives of Disease in Childhood. 2010 Sep 1;95(9):662-7.
13. Shann F. Nonspecific effects of vaccines and the reduction of mortality in children. Clinical Therapeutics. 2013 Feb 1;35(2):109-14.
14. Nemes E, Geldenhuys H, Rozot V, Rutkowski KT, Ratangee F, Bilek N, et al. Prevention of M. tuberculosis infection with H4: IC31 vaccine or BCG revaccination. New England Journal of Medicine. 2018 Jul 12;379(2):138-49.
15. Aaby P, Martins CL, Garly ML, Andersen A, Fisker AB, Claesson MH, et al. Measles vaccination in the presence or absence of maternal measles antibody: impact on child survival. Clinical Infectious Diseases. 2014 Aug 15;59(4):484-92.
16. Clark IA, Allison AC, Cox FE. Protection of mice against Babesia, and Plasmodium with BCG. Nature. 1976 Jan;259(5541):309-11.
17. de Bree LC, Koeken VA, Joosten LA, Aaby P, Benn CS, van Crevel R, et al. Non-specific effects of vaccines: Current evidence and potential implications. InSeminars in Immunology 2018 Oct 1; 39:35-43.
18. Roth A, Gustafson P, Nhaga A, Djana Q, Poulsen A, Garly ML, et al. BCG vaccination scar associated with better childhood survival in Guinea-Bissau. International Journal of Epidemiology. 2005 Jun 1;34(3):540-7.
19. Higgins JP, Soares-Weiser K, López-López JA, Kakourou A, Chaplin K, Christensen H, et al. Association of BCG, DTP, and measles containing vaccines with childhood mortality: systematic review. BMJ. 2016 Oct 13;355.
20. Stensballe LG, Nante E, Jensen IP, Kofoed PE, Poulsen A, Jensen H, et al. Acute lower respiratory tract infections and respiratory syncytial virus in infants in Guinea-Bissau: a beneficial effect of BCG vaccination for girls: community based case–control study. Vaccine. 2005 Jan 26;23(10):1251-7.
21. Wardhana DE, Sultana A, Mandang VV, Jim E. The efficacy of Bacillus Calmette-Guerin vaccinations for the prevention of acute upper respiratory tract infection in the elderly. Acta Med Indones. 2011 Jul;43(3):185-90.
22. Ohrui T, Nakayama K, Fukushima T, Chiba H, Sasaki H. Prevention of elderly pneumonia by pneumococcal, influenza and BCG vaccinations. Nihon Ronen Igakkai zasshi. Japanese Journal of Geriatrics. 2005 Jan 1;42(1):34-6.
23. Aaby P, Benn CS. Saving lives by training innate immunity with bacille Calmette-Guerin vaccine. Proceedings of the National Academy of Sciences. 2012 Oct 23;109(43):17317-8.
24. Biering-Sørensen S, Aaby P, Napirna BM, Roth A, Ravn H, Rodrigues A, et al. Small randomized trial among low– birth-weight children receiving Bacillus Calmette-Guérin vaccination at first health center contact. The Pediatric Infectious Disease Journal. 2012 Mar 1;31(3):306-8.
25. Aaby P, Roth A, Ravn H, Napirna BM, Rodrigues A, Lisse IM, et al. Randomized trial of BCG vaccination at birth to low-birth-weight children: beneficial nonspecific effects in the neonatal period?. Journal of Infectious Diseases. 2011 Jul 15;204(2):245-52.
26. Ponnighaus JM, Msosa E, Gruer PJ, Liomba NG, Fine PE, Sterne JA, et al. Efficacy of BCG vaccine against leprosy and tuberculosis in northern Malawi. The Lancet. 1992 Mar 14;339(8794):636-9.
27. Garly ML, Martins CL, Balé C, Baldé MA, Hedegaard KL, Gustafson P, et al. BCG scar and positive tuberculin reaction associated with reduced child mortality in West Africa: a non-specific beneficial effect of BCG?. Vaccine. 2003 Jun 20;21(21-22):2782-90.
28. Goodridge HS, Ahmed SS, Curtis N, Kollmann TR, Levy O, Netea MG, et al. Harnessing the beneficial heterologous effects of vaccination. Nature Reviews Immunology. 2016 Jun;16(6):392-400.
29. Ota MO, Vekemans J, Schlegel-Haueter SE, Fielding K, Sanneh M, Kidd M, et al. Influence of Mycobacterium bovis bacillus Calmette-Guerin on antibody and cytokine responses to human neonatal vaccination. The Journal of Immunology. 2002 Jan 15;168(2):919-25.
30. Ritz N, Mui M, Balloch A, Curtis N. Non-specific effect of Bacille Calmette-Guerin vaccine on the immune response to routine immunisations. Vaccine. 2013 Jun 26;31(30):3098-103.
31. Bree CD, Crevel RV, Netea MG. H4: IC31 Vaccine or BCG revaccination for tuberculosis. New England Journal of Medicine. 2018;379:1969.
32. Kleinnijenhuis J, Quintin J, Preijers F, Joosten LA, Ifrim DC, Saeed S, et al. Bacille Calmette-Guerin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proceedings of the National Academy of Sciences. 2012 Oct 23;109(43):17537-42.
33. Takeda K, Akira S. TLR signaling pathways. InSeminars in Immunology 2004 Feb 1;16(1):3-9.
34. Li L, Qiao D, Zhang X, Liu Z, Wu C. The immune responses of central and effector memory BCG-specific CD4+ T cells in BCG-vaccinated PPD+ donors were modulated by Treg cells. Immunobiology. 2011 Apr 1;216(4):477-84.
35. Soares AP, Kwong Chung CK, Choice T, Hughes EJ, Jacobs G, van Rensburg EJ, et al. Longitudinal changes in CD4+ T-cell memory responses induced by BCG vaccination of newborns. The Journal of Infectious Diseases. 2013 Apr 1;207(7):1084-94.
36. Ovchinnikova OA, Berge N, Kang C, Urien C, Ketelhuth DF, Pottier J, et al. Mycobacterium bovis BCG killed by extended freeze-drying induces an immunoregulatory profile and protects against atherosclerosis. Journal of Internal Medicine. 2014 Jan;275(1):49-58.
37. Coleman MM, Keane J, Mills KH. Tregs and BCG— dangerous liaisons in TB. Journal of Leukocyte Biology. 2010 Dec;88(6):1067-9.
38. Sebina I, Cliff JM, Smith SG, Nogaro S, Webb EL, Riley EM, et al. Long-lived memory B-cell responses following BCG vaccination. PloS One. 2012 Dec 11;7(12):e51381.
39. Saint F, Patard JJ, Maille P, Soyeux P, Hoznek A, Salomon L, et al. Prognostic value of at helper 1 urinary cytokine response after intravesical bacillus Calmette- Guérin treatment for superficial bladder cancer. The Journal of Urology. 2002 Jan;167(1):364-7.
40. Eyerich S, Eyerich K, Pennino D, Carbone T, Nasorri F, Pallotta S, et al. Th22 cells represent a distinct human T cell subset involved in epidermal immunity and remodeling. The Journal of Clinical Investigation. 2009 Dec 1;119(12):3573-85.
41. Brandau S, Suttmann H, Flad HD, Jocham D, Böhle A. Killing of Fas ligand-resistant renal carcinoma cells by interleukin-2-and BCG-activated effector cells. Cancer Immunology, Immunotherapy. 2000 Aug 1;49(7):369-76.
42. Brandau S, Riemensberger J, Jacobsen M, Kemp D, Zhao W, Zhao X, et al. NK cells are essential for effective BCG immunotherapy. International Journal of Cancer. 2001 Jun 1;92(5):697-702.
43. Riemensberger J, Böhle A, Brandau S. IFN-gamma and IL-12 but not IL-10 are required for local tumour surveillance in a syngeneic model of orthotopic bladder cancer. Clinical & Experimental Immunology. 2002 Jan;127(1):20-6.
44. Reactome icons library. https://reactome.org/iconlib. (accessed 24 August 2020).
45. Master SS, Rampini SK, Davis AS, Keller C, Ehlers S, Springer B, et al. Mycobacterium tuberculosis prevents inflammasome activation. Cell host & microbe. 2008 Apr 17;3(4):224-32.
46. Read SA, Obeid S, Ahlenstiel C, Ahlenstiel G. The role of zinc in antiviral immunity. Advances in nutrition. 2019 Jul 1;10(4):696-710.
47. Johansen P, Fettelschoss A, Amstutz B, Selchow P, Waeckerle-Men Y, Keller P, et al. Relief from Zmp1- mediated arrest of phagosome maturation is associated with facilitated presentation and enhanced immunogenicity of mycobacterial antigens. Clinical and Vaccine Immunology. 2011 Jun 1;18(6):907-13.
48. Mueller SN, Rouse BT. Immune responses to viruses. Clinical Immunology. 2008:421.
49. García-Sastre A, Biron CA. Type 1 interferons and the virus-host relationship: a lesson in detente. Science. 2006 May 12;312(5775):879-82.
50. Kawai T, Akira S. Innate immune recognition of viral infection. Nature Immunology. 2006 Feb;7(2):131-7.
51. Chow KT, Gale Jr M, Loo YM. RIG-I and other RNA sensors in antiviral immunity. Annual Review of Immunology. 2018 Apr 26;36:667-94.
52. Leonard JN, Ghirlando R, Askins J, Bell JK, Margulies DH, Davies DR, et al. The TLR3 signaling complex forms by cooperative receptor dimerization. Proceedings of the National Academy of Sciences. 2008 Jan 8;105(1):258-63.
53. Tissari J, Sirén J, Meri S, Julkunen I, Matikainen S. IFN-α enhances TLR3-mediated antiviral cytokine expression in human endothelial and epithelial cells by upregulating TLR3 expression. The Journal of Immunology. 2005 Apr 1;174(7):4289-94.
54. V’kovski P, Kratzel A, Steiner S, Stalder H, Thiel V. Coronavirus biology and replication: implications for SARS-CoV-2. Nature Reviews Microbiology. 2020 Oct 28:1-6.
55. Astuti I. Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2): An overview of viral structure and host response. Diabetes & Metabolic Syndrome: Clinical Research & Reviews. 2020 Apr 18.
56. Pobezinskaya YL, Kim YS, Choksi S, Morgan MJ, Li T, Liu C, et al. The function of TRADD in signaling through tumor necrosis factor receptor 1 and TRIF-dependent Toll- like receptors. Nature Immunology. 2008 Sep;9(9):1047-54.
57. Ermolaeva MA, Michallet MC, Papadopoulou N, Utermöhlen O, Kranidioti K, Kollias G, et al. Function of TRADD in tumor necrosis factor receptor 1 signaling and in TRIF-dependent inflammatory responses. Nature Immunology. 2008 Sep;9(9):1037-46.
58. Jensen S, Thomsen AR. Sensing of RNA viruses: a review of innate immune receptors involved in recognizing RNA virus invasion. Journal of Virology. 2012 Mar 15;86(6):2900-10.
59. Li G, Fan Y, Lai Y, Han T, Li Z, Zhou P, et al. Coronavirus infections and immune responses. Journal of Medical Virology. 2020 Apr;92(4):424-32.
60. Arts RJ, Moorlag SJ, Novakovic B, Li Y, Wang SY, Oosting M, et al. BCG vaccination protects against experimental viral infection in humans through the induction of cytokines associated with trained immunity. Cell Host & Microbe. 2018 Jan 10;23(1):89-100.
61. COVID-19 coronavirus pandemic. https://www. worldometers.info/coronavirus/ (accessed 28 August 2020).