Editorial
Most of what we know about oral disease is measured in bulk. Saliva is pooled, plaque is scraped from multiple surfaces and lumped together, and gingival samples are homogenized before downstream analyses. The resulting biomarker or transcriptome profiles average over compartments that differ by cell type, by oxygen tension, by microbial neighborhood, and by mechanical properties. Yet dental plaque is a micron-structured community, periodontal tissue is layered into epithelial, mesenchymal, and vascular compartments, and endodontic and peri-implant biofilms form microcolonies with gradients spanning tens of micrometers. In contrast to traditional technologies, spatial transcriptomics can provide information on the location and organization of microbial diversity and host gene expression signals, helping us accurately understand the biology of oral tissues. Research in oral health should adopt these methods now, for three reasons.
First, spatial methods have repeatedly recovered disease-relevant signals that bulk approaches have averaged away in tissues outside the oral cavity. A 2026 single-cell spatial atlas of human skin used multiplexed error-robust fluorescence in situ hybridization (MERFISH) resolved a perivascular neighborhood with shared alterations across atopic dermatitis, psoriasis, and hidradenitis suppurativa, and identified a gene expression program maintaining perivascular fibroblasts that bulk RNA sequencing had not isolated [1]. In chronic active multiple sclerosis, spatial transcriptomics of subcortical lesions resolved cell-type-specific niche programs at the lesion edge, including microglia and astrocyte states, that whole-lesion bulk averaging had hidden [2]. In each case, the disease biology was organized at a scale finer than the usual bulk measurement, and averaging over that organization erased the signal of interest. Spatial transcriptomics technologies such as Visium HD (10x Genomics, Pleasanton, CA, USA), Xenium (10x Genomics) and MERFISH (Vizgen, Cambridge, MA, USA) are now commercially available for overcoming this barrier, making it possible to assess panels of several thousand genes at sub-micrometer spatial resolution [3].
Second, the structures that drive oral disease specifically are organized at micron scale, not as homogeneous tissue. Supragingival plaque is not a uniform biofilm but rather a collection of highly organized “hedgehog” and “corncob” structures, with Corynebacterium filaments anchoring a scaffold, Streptococcus and Neisseriaceae at the aerobic periphery, and Fusobacterium and Leptotrichia in a microaerobic annulus, just to name a few key taxa [4,5]. The architecture is consistent with steep oxygen and metabolic gradients across tens of micrometers. The imaging methods that revealed this organization: combinatorial-label spectral-imaging fluorescence in situ hybridization (FISH) and its successors, are multiplexed to profile fifty oral genera in a single specimen through imaging [6]. Additionally, close relatives within the same genus separate cleanly across tongue dorsum, buccal mucosa, and supragingival and subgingival habitats, so that taxa traditionally considered to be “generalists” resolve into niche-specific subtypes when sampled at the right scale [7]. Similarly, in peri-implant tissue, preliminary data indicate that healthy biofilms appear as mixed taxonomic patches of five to twenty-five micrometers, while peri-implantitis is marked by monolithic single-genus patches of fifty to one hundred micrometers with sharp boundaries [8]. Microscopy has resolved this architecture, but the underlying gene-expression programs at the same scale have not been measured. The same applies for host tissue. The junctional epithelium, sulcular epithelium, gingival rete ridges, periodontal ligament, and pulp–dentin interface each carry distinct transcriptional programs, and detailed spatial data of these compartments at micron scale are still lacking.
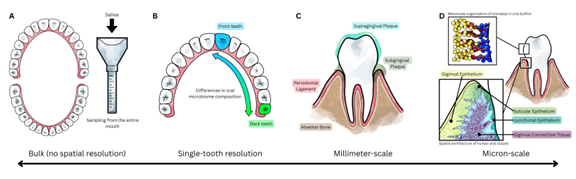
Figure 1. Scales of spatial transcriptomics in oral health. (A) Bulk measurement returns one signal that is averages over a whole tissue compartment, such as the entire oral cavity. (B) Analyses at the single-tooth resolution, while coarse, can identify differences and gradients between the anterior and posterior teeth. (C) Millimeter-scale spatial stratification into supragingival plaque, subgingival plaque, gingival epithelium, periodontal ligament, and bone. (D) Micron?scale spatial transcriptomics will enable the exploration of oral tissues at unprecedented resolution, from resolving individual microbial taxa within highly organized biofilm (upper), to profiling distinct host cell states across the gingival epithelium and connective tissue (lower).
Third, even at very coarse spatial resolution, the application of “spatial transcriptomics” has already uncovered biologically relevant signals. The first spatial transcriptomic study of a periodontitis biopsy used Visium at fifty-five-micrometer spots and separated epithelium, inflamed connective tissue, and non-inflamed connective tissue, recovering strong plasma-cell signatures that pooled gingival extracts had obscured [9]. A 2024 critical review of oral and craniofacial applications reported that markers of periodontitis susceptibility localize specifically to junctional epithelium rather than to gingiva as a whole [10]. Other spatially resolved analyses uncovered niches of disease-relevant cell states, such as sulcular and junctional keratinocyte states in compartmentalized immune niches across diseased sites[11], and a pro-inflammatory fibroblast subset driving cytokine gradients [12]. In caries, direct in situ imaging has shown that niches of acidogenic microbes occupy precise positions on the tooth surface [13], and that cariogenic metabolic activity concentrates in a small minority of taxa that bulk profiling would miss [14]. These promising early findings suggest that further exploration of oral tissues using spatial transcriptomics may hold the key to developing novel diagnostics and therapeutics for oral diseases.
In the near term, the application of spatial transcriptomic tools may help answer three clinically relevant questions. In caries, understanding the role that biofilm organization plays in caries progression may further the development of caries diagnostics and therapeutics. In endodontics, better understanding the pulp’s reaction to infection may help distinguish reversible pulpitis from irreversible pulpitis: a diagnostic decision that currently relies on clinical heuristics without a molecular basis [15]. Similarly, in periodontitis, mapping the junctional-epithelium niche where immune activation leads to alveolar bone loss can help distinguishing aggressive from chronic disease and propose new avenues for treatment. From the tooth surface to the pulp to the periodontium, spatially complex structures govern the balance between oral health versus oral disease, and resolving these spatial signals may hold the key towards solving the challenges in oral health.
References
2. Lerma-Martin C, Badia-I-Mompel P, Ramirez Flores RO, Sekol P, Schäfer PSL, Riedl CJ, et al. Cell type mapping reveals tissue niches and interactions in subcortical multiple sclerosis lesions. Nat Neurosci. 2024 Dec;27(12):2354–65.
3. Lim HJ, Wang Y, Buzdin A, Li X. A practical guide for choosing an optimal spatial transcriptomics technology from seven major commercially available options. BMC Genomics. 2025 Jan 20;26(1):47.
4. Borisy GG, Valm AM. Spatial scale in analysis of the dental plaque microbiome. Periodontol 2000. 2021 Jun;86(1):97–112.
5. Mark Welch JL, Rossetti BJ, Rieken CW, Dewhirst FE, Borisy GG. Biogeography of a human oral microbiome at the micron scale. Proc Natl Acad Sci U S A. 2016 Feb 9;113(6):E791–800.
6. Valm AM, Mark Welch JL, Rieken CW, Hasegawa Y, Sogin ML, Oldenbourg R, et al. Systems-level analysis of microbial community organization through combinatorial labeling and spectral imaging. Proc Natl Acad Sci U S A. 2011 Mar 8;108(10):4152–7.
7. Mark Welch JL, Dewhirst FE, Borisy GG. Biogeography of the Oral Microbiome: The Site-Specialist Hypothesis. Annu Rev Microbiol. 2019 Sep 8;73:335–58.
8. Grodner B, Wu DT, Hahm S, Takayasu L, Wen N, Kim DM, et al. Microscale Spatial Dysbiosis in Oral biofilms Associated with Disease. bioRxiv. 2024 Jul 24:2024–07.
9. Lundmark A, Gerasimcik N, Båge T, Jemt A, Mollbrink A, Salmén F, et al. Gene expression profiling of periodontitis-affected gingival tissue by spatial transcriptomics. Sci Rep. 2018 Jun 19;8(1):9370.
10. Haller J, Abedi N, Hafedi A, Shehab O, Wietecha MS. Spatial Transcriptomics Unravel the Tissue Complexity of Oral Pathogenesis. J Dent Res. 2024 Dec;103(13):1331–9.
11. Easter QT, Fernandes Matuck B, Beldorati Stark G, Worth CL, Predeus AV, Fremin B, et al. Single-cell and spatially resolved interactomics of tooth-associated keratinocytes in periodontitis. Nat Commun. 2024 Jun 14;15(1):5016.
12. Caetano AJ, Redhead Y, Karim F, Dhami P, Kannambath S, Nuamah R, et al. Spatially resolved transcriptomics reveals pro-inflammatory fibroblast involved in lymphocyte recruitment through CXCL8 and CXCL10. Elife. 2023 Jan 17;12:e81525.
13. Kim D, Barraza JP, Arthur RA, Hara A, Lewis K, Liu Y, et al. Spatial mapping of polymicrobial communities reveals a precise biogeography associated with human dental caries. Proc Natl Acad Sci U S A. 2020 Jun 2;117(22):12375–86.
14. Naumann AA, Elmorr EM, Lamont EI, Hendrickson EL, Holmes T, Nelson TM, et al. DNA and RNA-based amplicon sequencing of paired supragingival and dentin lesion plaque in children with severe early childhood caries. Int J Oral Sci. 2026 Feb 24;18(1):19.
15. Mejàre IA, Axelsson S, Davidson T, Frisk F, Hakeberg M, Kvist T, et al. Diagnosis of the condition of the dental pulp: a systematic review. Int Endod J. 2012 Jul;45(7):597–613.