Abstract
Lung cancer is the leading cause of cancer-related death in men and women with a 5-year survival rate of 18%. The PI3K/Akt pathway plays a role in non-small cell lung cancer tumorigenesis and progression. FCH domain only 1(FCHO1) is overexpressed in lung cancer and promotes entry into mitosis and G1/S phase transition, leading to cancer cell growth. Our group has investigated the relationship between FCHO1 and the Akt signaling pathway, which plays an important role in lung cancer growth. Serine 570 of human FCHO1, a part of the Akt kinase motif, was known to be phosphorylated by Akt. To inhibit FCHO1 phosphorylation, we developed the FCHO1560-571 peptide. FCHO1560-571 peptide exhibits higher binding affinity to Akt and inhibits lung cancer cell proliferation. Moreover, the FCHO1560-571 peptide significantly decreased p-Akt, p-ERK and CDC2 levels. These results suggest that the FCHO1560-571 peptide inhibits cancer progression through the Akt/ERK/CDC2 pathway.
There are few studies investigating the function of FCHO1 in cancer cell growth. In this mini review, we summarize recent articles on the role of FCHO1 in lung cancer progression. The reviewed articles suggest that inhibition of FCHO1 activity through the regulation of Akt signaling could be developed as a novel lung cancer treatment.
Keywords
Lung cancer, Akt signaling pathway, FCHO1, Cell cycle, Midbody
Activation of Akt Promotes Lung Cancer Progression
Akt mediates growth factor-associated cell survival. Constitutive activation of Akt (phosphorylated Akt, p-Akt) occurs in several human cancers, including lung cancer, and may be associated with poor prognosis and chemotherapy and radiotherapy resistance [1-3] .
Akt, or protein kinase B, a serine/threonine protein kinase [4], is activated downstream from phosphatidylinositol 3-kinase (PI3K), by various growth factors, including insulin, insulin-like growth factor-I, and epidermal growth factor [5]. Activated Akt (phospho-Akt, p-Akt) is a strong promoter of cell survival because it antagonizes and inactivates various components of the apoptotic cascade such as proapoptotic Bad, caspase-9, and forkhead transcription factor family members [6,7]. The PI3K/Akt signaling pathway regulates the G1/S cell cycle transition by modulating the transcription of cell cycle proteins and suppressing cell cycle inhibitors [8]. A dysregulated cell cycle is often associated with increased tumorigenesis and accelerated tumor growth [9].
FCH Domain Only 1 (FCHO1) Promotes Lung Cancer Growth
Clathrin-mediated endocytosis is the major endocytic pathway by which eukaryotic cells internalize cell-surface cargo proteins and extracellular molecules, thereby enabling a broad range of biological processes, including cell signaling, nutrient and growth factor uptake, and cell fate and differentiation [10].
FCH domain only 1 (FCHO1) is a member of the Fer/CIP4 homology-Bin/amphiphysin/Rvs (F-BAR) protein family, which is a characterized by the presence of the F-BAR domain [11]. The F-BAR domain of FCHO1 is associated with membrane tubulation during early endocytosis [12].
FCHO1 with truncations or mutations in the F-BAR domain was isolated from patients with T cell lymphopenia. Because these mutations severely perturb T cell receptor endocytosis, FCHO1 affects T cell and receptor-mediated immune responses [13]. Silencing of FCHO1 causes severe developmental defects in zebrafish [14]. These data suggest that FCHO1 is involved in the regulation of cell growth and development.
Overexpression of FCHO1 affects mitosis in lung cancer [15], and FCHO1 plays an important role in the G1/S phase transition and may be related to cancer, immune responses, and diseases related to growth disorders.
However, there are few studies focusing on the relationship between FCHO1 and cancer [15,17]. Recently, we studied the effect of FCHO1 on lung cancer and found that FCHO1 is overexpressed in many cancer cells [15]. We showed that overexpression of FCHO1 promotes lung cancer cell growth by activating cell division [16]. It was confirmed that the size of lung tumors was decreased in response to direct delivery of the shRNA FCHO1 in a lung cancer mouse model. In addition, FCHO1 is overexpressed in lymph node metastases from primary lung tumors [17]. These results suggest that overexpression of FCHO1 plays a significant role in the growth and metastasis of lung cancer [15,17] (Figure 1).
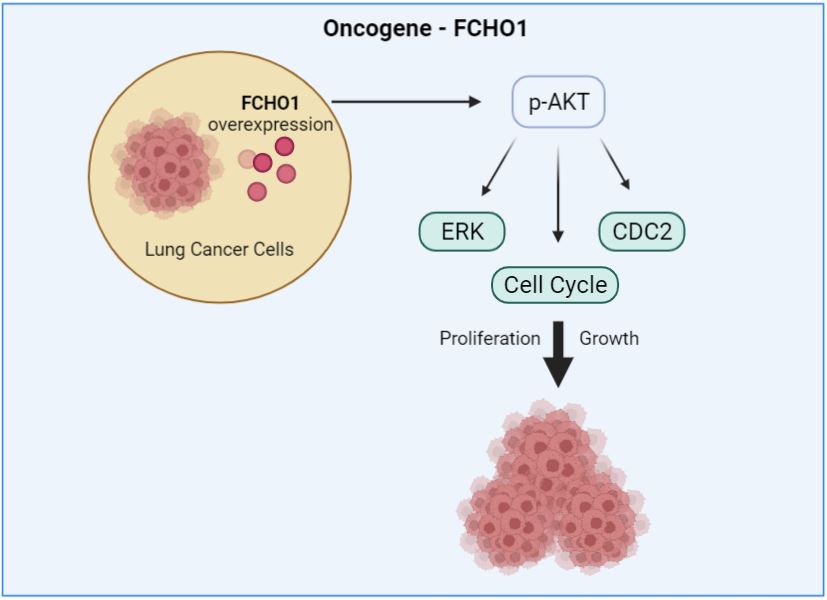
Figure 1: Effects of FCHO1 on lung cancer growth.
We investigated the relation between FCHO1 and the Akt signaling pathway, which plays an important role in lung cancer growth. Serine 570 of human FCHO1 is part of the kinase motif (RXRXXS) that is phosphorylated by Akt. We developed the FCHO1560-571 peptide, which inhibits Akt phosphorylation. The FCHO1 peptide containing the RXRXXS motif shows higher binding affinity to Akt, which suppresses lung cancer cell proliferation [16]. These results support that upregulation of FCHO1 in the lung promotes the progression of lung cancer by modulating cell-cycle progression.
FCHO1 Regulates Mitotic Proteins in Lung Cancer
Lung cancers develop the ability to bypass several cellcycle check points, and tumor cell proliferation is frequently associated with genetic or epigenetic alterations in key regulators of the cell cycle [18]. The PI3K/Akt/mTOR (mammalian target of rapamycin) signaling pathway is well characterized and plays essential roles in lung cancer cell proliferation and survival [19,20]. The PI3K/Akt pathway regulates the G1/S cell cycle transition by modulating the transcription of cell cycle proteins and suppressing cell cycle inhibitors [21].
Proteomics analysis after FCHO1 overexpression in lung cancer cells show that, FCHO1 regulates the expression of cell cycle-related proteins, particularly midbody proteins expressed in the G1/S phase [15,16].
Suppression of SMAD Family Member 4 (SMAD4) promotes metastasis of lung tumors by activating Akt [22]. A recent study showed that upregulation of SMAD4 inhibits Akt, thereby promoting lung cancer cell death induced by FCHO1 peptide. The FCHO1 peptide also induces G2/M cell cycle arrest and apoptosis by downregulating cell division control protein 2 (CDC2) and, Akt and inhibiting Extracellular signal-regulated kinase (ERK) phosphorylation [16]. Knockout of FCHO1 also affects mitochondrial electron-transport protein expression. An increase in mitochondrial electron-transport proteins occurs frequently during the G1/S phase transition [23].
FCHO1 was identified as a midbody protein by coimmunostaining with a midbody marker protein, although these results have not been published. Mitotic cell division is crucial for the proliferation, differentiation, and development of cells [24]. Failure to faithfully divide the genomic material, as well as cytoplasmic organelles and signaling molecules, leads to carcinogenesis and various developmental disorders [25]. Cytokinesis is a terminal stage of cell division, that results in the formation of two genetically identical daughter cells connected by an intercellular bridge containing the midbody [26]. Several major signaling pathways involved in cell proliferation, migration and cell fate have been localized to the midbody [27]. The midbody protein FCHO1 shRNA induces cell death by inhibiting lung cancer mitosis [15], suggesting that suppression of FCHO1 expression affects G1/S phase transition and could be developed as a therapeutic strategy for lung cancer [15].
Therapeutic Strategy Based on FCHO1 Function
Akt phosphorylation and cell-cycle progression are essential for cell growth [28]; therefore, Akt phosphorylation and G1/S phase regulators are targets for cancer treatment [29,30]. The role of FCHO1 in cancer and related signaling pathways has not been investigated to date, although data from our group suggest that FCHO1 overexpressed in cancer cells.
Evidence indicates that FCHO1 overexpression plays a role in lung cancer growth, which highlights the need to identify and develop novel targets to inhibit FCHO1 expression [15,16]. In addition, the involvement of FCHO1 in cell growth suggests that developing therapeutic agents, targeting FCHO1 could be an effective strategy for the treatment of various disease including cancer [14].
siRNAs or peptides capable of inhibiting FCHO1 expression could be used to regulate Akt downstream-signaling pathways by directly inhibiting Akt activity [15,16]. These results may lead to the development of lung cancer therapeutics targeting FCHO1.
FCHO1 shRNA induces lung cancer cell death by inhibiting Aurora kinase B (Aurora B) which plays an important role in mitosis [15,31]. Although many drugs that inhibit the activity of Aurora kinase B are being developed in clinical practice, there are many limitations [32,33]. The development of a new therapeutic agent capable of inhibiting the expression of FCHO1 and simultaneously inhibiting Aurora kinase B activity will be important for the treatment of lung cancer.
MicroRNAs (miRNAs) are another approach to regulate the expression of FCHO1[34]. MiR-99a and miR-5481 directly regulate the PI3K/Akt signaling pathway and inhibit cell proliferation and invasion of lung cancer cells [35-37]. miR-99a or miR-5481-mediated inhibition of FCHO1 may thus suppress the proliferation of lung cancer cells. The inhibition of FCHO1 expression and the ability to modulate the associated Akt and cell cycle signaling pathways could be important strategies leading to novel lung cancer therapeutics (Figure 2).
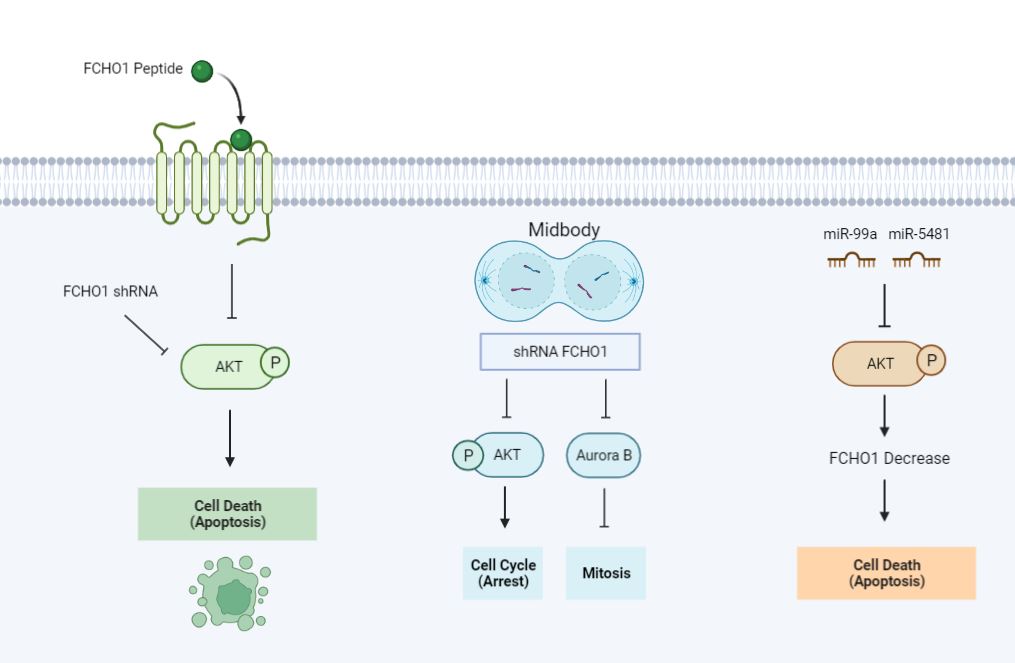
Figure 2: Schematic diagram of the development strategy of FCHO1 as a lung cancer treatment.
We recently showed that FCHO1 is a midbody protein that regulate mitosis [15]. Studies on the mechanism of action of FCHO1 as a mid-body protein involved in lung cancer cell death and progression will also be important for the development of new lung cancer therapeutics.
Conclusion
There are currently no studies investigating the role of FCHO1 in lung cancer progression. Work from our group showed that inhibition of FCHO1 expression and activity is a potential strategy for the treatment of lung cancer drugs capable of inhibiting FCHO1 directly affect the activity of Akt [16]. Recent work from our group showed that FCHO1 plays an oncogenic role in lung cancer [15]. Given the elevated level of FCHO1 expression and its role in lung cancer, therapeutic agents that modulate the Akt signaling pathway could be developed as targeted therapeutics for regulating FCHO1 in lung cancer.
Competing and Declaration of Interests
All authors have approved the manuscript and agreed with the submission. None of the authors have conflict of interest to declare.
Acknowledgements
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (NRF-2021R1A2C1007373).
References
2. Coffer PJ, Jin J, Woodgett JR. Protein kinase B (c-Akt): a multifunctional mediator of phosphatidylinositol 3-kinase activation. Biochem J. 1998;335 ( Pt 1):1-13.
3. Shah A, Swain WA, Richardson D, Edwards J, Stewart DJ, Richardson CM, et al. Phospho-akt expression is associated with a favorable outcome in non-small cell lung cancer. Clin Cancer Res. 2005;11(8):2930-6.
4. Song M, Bode AM, Dong Z, Lee MH. AKT as a Therapeutic Target for Cancer. Cancer Res. 2019;79(6):1019-31.
5. Liu P, Cheng H, Roberts TM, Zhao JJ. Targeting the phosphoinositide 3-kinase pathway in cancer. Nat Rev Drug Discov. 2009;8(8):627-44.
6. Brunet A, Bonni A, Zigmond MJ, Lin MZ, Juo P, Hu LS, et al. Akt promotes cell survival by phosphorylating and inhibiting a Forkhead transcription factor. Cell. 1999;96(6):857-68.
7. Osaki M, Oshimura M, Ito H. PI3K-Akt pathway: its functions and alterations in human cancer. Apoptosis. 2004;9(6):667-76.
8. Chang F, Lee JT, Navolanic PM, Steelman LS, Shelton JG, Blalock WL, et al. Involvement of PI3K/Akt pathway in cell cycle progression, apoptosis, and neoplastic transformation: a target for cancer chemotherapy. Leukemia. 2003;17(3):590-603.
9. Liang J, Slingerland JM. Multiple roles of the PI3K/PKB (Akt) pathway in cell cycle progression. Cell Cycle. 2003;2(4):339-45.
10. Mettlen M, Chen PH, Srinivasan S, Danuser G, Schmid SL. Regulation of Clathrin-Mediated Endocytosis. Annu Rev Biochem. 2018;87:871-96.
11. Henne WM, Kent HM, Ford MG, Hegde BG, Daumke O, Butler PJ, et al. Structure and analysis of FCHo2 F-BAR domain: a dimerizing and membrane recruitment module that effects membrane curvature. Structure. 2007;15(7):839-52.
12. Henne WM, Boucrot E, Meinecke M, Evergren E, Vallis Y, Mittal R, et al. FCHo proteins are nucleators of clathrin-mediated endocytosis. Science. 2010;328(5983):1281-4.
13. Lyszkiewicz M, Zietara N, Frey L, Pannicke U, Stern M, Liu Y, et al. Human FCHO1 deficiency reveals role for clathrin-mediated endocytosis in development and function of T cells. Nat Commun. 2020;11(1):1031.
14. Umasankar PK, Sanker S, Thieman JR, Chakraborty S, Wendland B, Tsang M, et al. Distinct and separable activities of the endocytic clathrin-coat components Fcho1/2 and AP-2 in developmental patterning. Nat Cell Biol. 2012;14(5):488-501.
15. Park S, Lee AY, Cho KC, Jung JH, Hong SH, Kim S, et al. FCH domain only 1 (FCHo1), a potential new biomarker for lung cancer. Cancer Gene Ther. 2021.
16. Park S, Hong Y, Lee S, Lee AY, Tran Q, Lee H, et al. FCHO1(560- 571) peptide, a PKB kinase motif, inhibits tumor progression. Biochem Biophys Res Commun. 2020;528(3):478-84.
17. Chen Z, Xiong S, Li J, Ou L, Li C, Tao J, et al. DNA methylation markers that correlate with occult lymph node metastases of nonsmall cell lung cancer and a preliminary prediction model. Transl Lung Cancer Res. 2020;9(2):280-7.
18. Eymin B, Gazzeri S. Role of cell cycle regulators in lung carcinogenesis. Cell Adh Migr. 2010;4(1):114-23.
19. Cheng H, Shcherba M, Pendurti G, Liang Y, Piperdi B, Perez- Soler R. Targeting the PI3K/AKT/mTOR pathway: potential for lung cancer treatment. Lung Cancer Manag. 2014;3(1):67-75.
20. Porta C, Paglino C, Mosca A. Targeting PI3K/Akt/mTOR Signaling in Cancer. Front Oncol. 2014;4:64.
21. Fekete M, Santiskulvong C, Eng C, Dorigo O. Effect of PI3K/Akt pathway inhibition-mediated G1 arrest on chemosensitization in ovarian cancer cells. Anticancer Res. 2012;32(2):445-52.
22. Liu J, Cho SN, Akkanti B, Jin N, Mao J, Long W, et al. ErbB2 Pathway Activation upon Smad4 Loss Promotes Lung Tumor Growth and Metastasis. Cell Rep. 2015;10(9):1599-613.
23. da Veiga Moreira J, Peres S, Steyaert JM, Bigan E, Pauleve L, Nogueira ML, et al. Cell cycle progression is regulated by intertwined redox oscillators. Theor Biol Med Model. 2015;12:10.
24. Dionne LK, Wang XJ, Prekeris R. Midbody: from cellular junk to regulator of cell polarity and cell fate. Curr Opin Cell Biol. 2015;35:51-8.
25. Pollard TD. Mechanics of cytokinesis in eukaryotes. Curr Opin Cell Biol. 2010;22(1):50-6.
26. Antanaviciute I, Gibieza P, Prekeris R, Skeberdis VA. Midbody: From the Regulator of Cytokinesis to Postmitotic Signaling Organelle. Medicina (Kaunas). 2018;54(4).
27. Chen CT, Ettinger AW, Huttner WB, Doxsey SJ. Resurrecting remnants: the lives of post-mitotic midbodies. Trends Cell Biol. 2013;23(3):118-28.
28. Pene F, Claessens YE, Muller O, Viguie F, Mayeux P, Dreyfus F, et al. Role of the phosphatidylinositol 3-kinase/Akt and mTOR/P70S6-kinase pathways in the proliferation and apoptosis in multiple myeloma. Oncogene. 2002;21(43):6587-97.
29. Shtivelman E, Sussman J, Stokoe D. A role for PI 3-kinase and PKB activity in the G2/M phase of the cell cycle. Curr Biol. 2002;12(11):919-24.
30. Kandel ES, Skeen J, Majewski N, Di Cristofano A, Pandolfi PP, Feliciano CS, et al. Activation of Akt/protein kinase B overcomes a G(2)/m cell cycle checkpoint induced by DNA damage. Mol Cell Biol. 2002;22(22):7831-41.
31. Ahmed A, Shamsi A, Mohammad T, Hasan GM, Islam A, Hassan MI. Aurora B kinase: a potential drug target for cancer therapy. J Cancer Res Clin Oncol. 2021;147(8):2187-98.
32. Yan M, Wang C, He B, Yang M, Tong M, Long Z, et al. Aurora-A Kinase: A Potent Oncogene and Target for Cancer Therapy. Med Res Rev. 2016;36(6):1036-79.
33. Borah NA, Reddy MM. Aurora Kinase B Inhibition: A Potential Therapeutic Strategy for Cancer. Molecules. 2021;26(7).
34. Bueno MJ, Perez de Castro I, Malumbres M. Control of cell proliferation pathways by microRNAs. Cell Cycle. 2008;7(20):3143- 8.
35. Yu SH, Zhang CL, Dong FS, Zhang YM. miR-99a suppresses the metastasis of human non-small cell lung cancer cells by targeting AKT1 signaling pathway. J Cell Biochem. 2015;116(2):268-76.
36. Liu C, Yang H, Xu Z, Li D, Zhou M, Xiao K, et al. microRNA-548l is involved in the migration and invasion of non-small cell lung cancer by targeting the AKT1 signaling pathway. J Cancer Res Clin Oncol. 2015;141(3):431-41.
37. Mei LL, Qiu YT, Huang MB, Wang WJ, Bai J, Shi ZZ. MiR-99a suppresses proliferation, migration and invasion of esophageal squamous cell carcinoma cells through inhibiting the IGF1R signaling pathway. Cancer Biomark. 2017;20(4):527-37.