Abstract
MicroRNAs (miRNAs) have emerged as central regulators of antitumor immunity, influencing immune checkpoint signaling, immune cell differentiation, and intercellular communication within the tumor microenvironment. Although extensive preclinical evidence supports the immunomodulatory potential of miRNAs, their clinical translation remains challenging due to context dependence, network redundancy, delivery limitations, and safety concerns. Building upon recent advances in single-cell and spatial profiling, extracellular vesicle biology, and miRNA delivery technologies, this invited commentary critically examines how these developments extend current understanding of miRNA-mediated immune regulation in cancer. We highlight key conceptual and translational bottlenecks that continue to hinder therapeutic implementation and discuss why miRNAs are unlikely to achieve optimal efficacy as standalone immunotherapies. Instead, emerging evidence supports their greatest near-term clinical impact as adjunctive modulators and dynamic biomarkers within existing immunotherapeutic frameworks. By integrating mechanistic insights with translational perspectives, this commentary outlines future directions emphasizing cell-specific targeting, network-informed strategies, and biomarker-guided integration, providing a forward-looking framework for harnessing miRNAs in next-generation cancer immunotherapy.
Keywords
microRNAs, Cancer immunotherapy, Tumor microenvironment, Immune checkpoint regulation, Extracellular vesicles, Biomarker-guided therapy, Immune reprogramming
Introduction
Recent years have firmly established microRNAs (miRNAs) as central master regulators of antitumor immunity, influencing immune checkpoint signaling, immune cell differentiation, and intercellular communication within the tumor microenvironment [1,2]. Over the past decade, extensive experimental and translational studies have established that miRNA-mediated post-transcriptional regulation influences virtually every stage of the cancer immunity cycle, from antigen presentation and T-cell priming to immune evasion and therapeutic resistance. In our recently published review, we provided a comprehensive synthesis of the molecular mechanisms by which miRNAs modulate immune responses in cancer. The present commentary is not a reiteration of that review but a forward-looking conceptual extension that integrates recent advances, emerging translational challenges, and future directions shaping the clinical development of miRNA-based immunomodulation with particular emphasis on immune checkpoints, extracellular vesicle (EV)-mediated communication, and immune cell reprogramming. For instance, several miRNAs have been shown to directly regulate immune checkpoint expressions, including miR-138-5p and miR-34a, both of which suppress PD-L1 expression in tumor cells and thereby enhance antitumor T-cell responses.
Building upon this foundation, the present commentary aims to move beyond a descriptive overview and instead critically examine how recent discoveries, unresolved challenges, and emerging technologies are reshaping the landscape of miRNA-based cancer immunomodulation. In particular, advances in single-cell sequencing, spatial transcriptomics, and EV biology have revealed an unexpected level of miRNA heterogeneity and context dependence that complicates their therapeutic exploitation [3,4]. At the same time, innovative delivery strategies, engineered EVs, and combinatorial immunotherapeutic approaches are renewing interest in miRNAs as viable clinical tools [5]. Despite compelling preclinical evidence, the clinical translation of miRNA-based therapies has lagged expectations. Factors such as off-target effects, immune-related toxicity, limited tissue specificity, functional variability of individual miRNAs across immune contexts continues to complicate their therapeutic application [6]. These challenges accentuate the need for a more nuanced understanding of miRNA regulatory networks rather than single-miRNA paradigms.
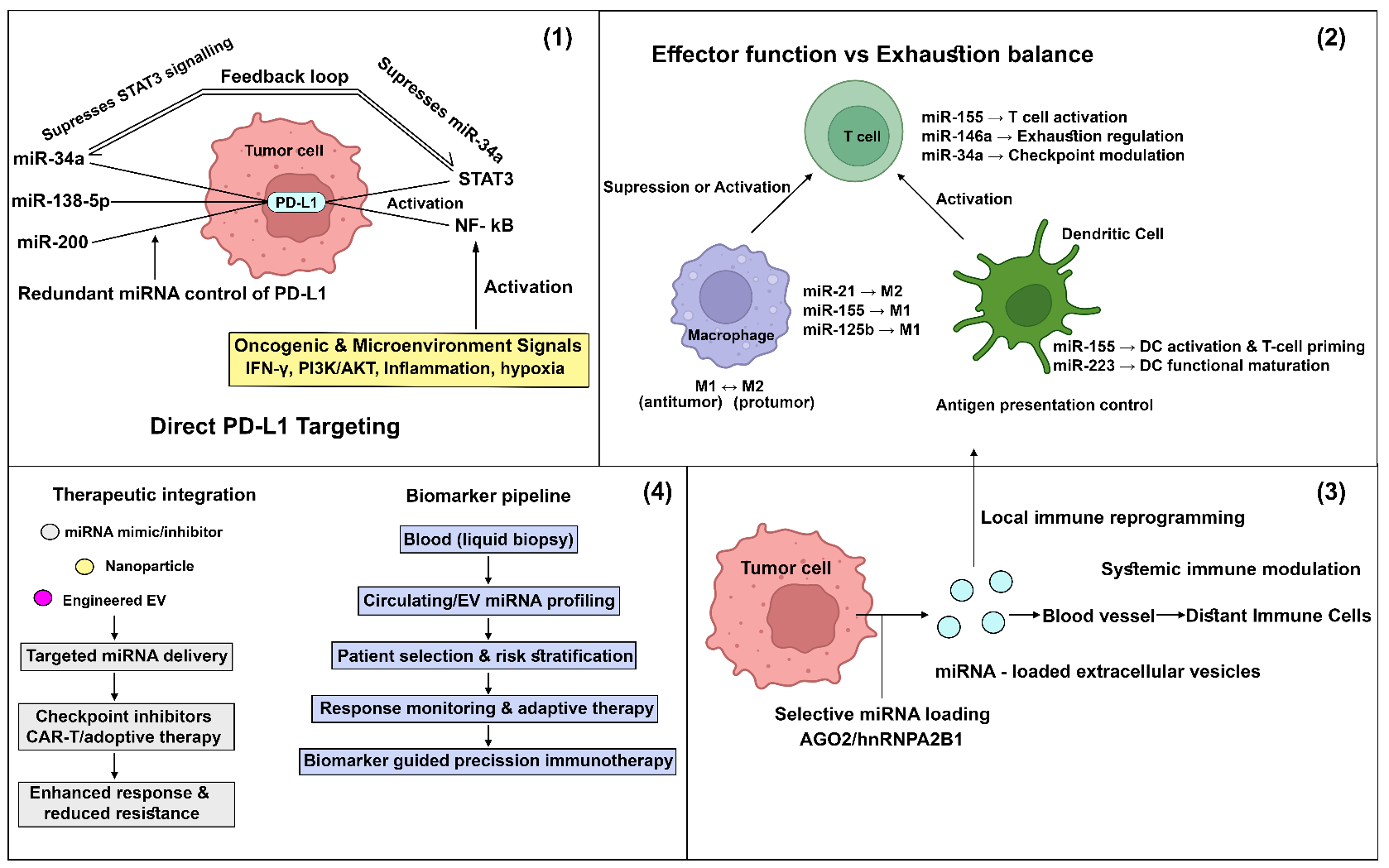
In this invited commentary, we discuss recent advances that extend the findings of the focal review, analyze key conceptual and translational bottlenecks, and highlight future directions that may enable the rational reprogramming of antitumor immunity through miRNA-based strategies. By integrating emerging mechanistic insights with therapeutic perspectives, we aim to provide a forward-looking framework for researchers and clinicians seeking to harness miRNAs for next-generation cancer immunotherapy. The conceptual framework of microRNA-mediated reprogramming of antitumor immunity and its translational integration is summarized in Figure 1.
Recent Advances Extending the Focal Review
Rapid methodological and conceptual advances over the past few years have substantially expanded our understanding of how microRNAs (miRNAs) regulate antitumor immunity beyond the canonical pathways described in earlier reviews. The integration of single-cell RNA sequencing and spatial transcriptomics has been particularly transformative, revealing pronounced heterogeneity in miRNA expression across tumor cells, stromal compartments, and immune populations [7]. These approaches demonstrate that miRNA-mediated immune regulation is highly cell-type, state, and microenvironment-dependent, challenging earlier assumptions that individual miRNAs exert uniform immunological effects across tumors.
Recent studies further indicate that immune checkpoint regulation is frequently governed by coordinated miRNA networks rather than single miRNAs. For example, PD-L1 expression is now recognized to be controlled by multiple miRNAs converging on shared signaling axes. miR-138-5p has been shown to directly bind the 3′UTR of PD-L1 and suppress its expression in several tumor models, thereby restoring T-cell activity. Similarly, miR-34a, one of the most extensively studied tumor-suppressive miRNAs, directly targets PD-L1 and enhances cytotoxic T-cell–mediated immune responses. Beyond individual examples, these findings illustrate that immune checkpoint regulation is governed by coordinated miRNA networks involving PI3K–AKT, JAK–STAT, and NF-κB pathways [1,6,8]. This multilayered regulation provides tumors with functional redundancy, helping to explain why therapeutic modulation of individual miRNAs often results in limited or transient immunotherapeutic benefit. These findings emphasize the importance of adopting network-based approaches to miRNA targeting.
Advances in extracellular vesicle (EV) biology have further reshaped the field. Tumor-derived EVs are no longer viewed as passive carriers but as actively regulated signaling entities. Selective miRNA loading is governed by RNA-binding proteins such as hnRNPA2B1 and AGO2, as well as sequence-specific motifs [2,9,10]. EV-associated miRNAs have been shown to exert durable immunomodulatory effects on recipient immune cells, including macrophages, dendritic cells, and natural killer cells, even at low copy numbers [11–13]. Importantly, emerging evidence suggests that EV-mediated miRNA transfer may contribute to systemic immune reprogramming, extending tumor-driven immunosuppression beyond the local microenvironment. Parallel progress in miRNA delivery technologies has renewed translational interest in miRNA-based immunomodulation. Engineered nanoparticles, chemically stabilized oligonucleotides, and bioengineered EVs have demonstrated improved stability, biodistribution, and cellular uptake in preclinical models [14,15]. Of particular relevance, immune cell–targeted delivery strategies have shown promise in selectively modulating macrophages or dendritic cells, thereby minimizing off-target effects and immune-related toxicity [16]. These advances suggest that long-standing delivery barriers may be partially overcome through rational design and cell-specific targeting.
In addition to their therapeutic potential, miRNAs are increasingly recognized as dynamic biomarkers of immunotherapy response. Circulating EV-associated miRNAs and immune cell–specific miRNA signatures have been correlated with clinical outcomes in patients receiving immune checkpoint inhibitors [2]. Unlike static genomic alterations, miRNA profiles can reflect real-time immune states and therapy-induced adaptations, positioning them as attractive candidates for monitoring treatment response and resistance. These advances extend the scope of the focal review by revealing miRNA-mediated immune regulation as a context-dependent, spatially organized, and dynamically regulated process. This evolving understanding marks a transition from descriptive cataloguing toward mechanistic integration and translational refinement, laying the groundwork for more precise and effective miRNA-based immunotherapeutic strategies.
A representative example of network-level miRNA regulation can be observed in the control of immune checkpoint expression, particularly PD-L1. Rather than being governed by a single miRNA, PD-L1 expression is regulated by interconnected miRNA modules involving miR-34a, miR-200 family members, and miR-138, which collectively target PD-L1 transcripts and associated signaling pathways [17,18]. These miRNAs are themselves embedded within broader regulatory circuits involving transcription factors such as STAT3, MYC, and NF-κB. For instance, STAT3 activation can suppress tumour-suppressive miRNAs including miR-34a [19–21], thereby indirectly promoting PD-L1 expression, while miR-34a can in turn inhibit signaling intermediates that sustain STAT3 activation, forming a feedback regulatory loop. Such feed-forward and feedback interactions create a robust regulatory network that stabilizes immune checkpoint expression under inflammatory or oncogenic conditions [18,22]. Systems-level perturbation of this network through simultaneous modulation of multiple miRNAs or upstream signaling nodes has been proposed as a more effective strategy than single-miRNA targeting. This network-oriented perspective highlights how miRNA–transcription factor cross-regulation and pathway redundancy shape immune evasion and therapeutic response in the tumor microenvironment [23,24].
Unresolved Challenges and Conceptual Gaps
Despite substantial progress, several conceptual, technical, and translational challenges continue to limit the effective deployment of microRNA (miRNA)-based strategies for reprogramming antitumor immunity. A central challenge lies in the cell- and state-specific behavior of miRNAs, where their regulatory outputs vary across immune compartments and microenvironmental conditions. miRNAs that promote inflammatory or antitumor responses in dendritic cells or cytotoxic lymphocytes may simultaneously enhance immunosuppressive programs in macrophages or regulatory T cells under different conditions [25,26]. This functional plasticity complicates therapeutic design and highlights the risk of unintended immune modulation when miRNAs are manipulated systemically.
Another major challenge lies in the redundancy and robustness of miRNA regulatory networks. Immune checkpoint expression and immune cell differentiation are rarely controlled by a single miRNA; rather, they are governed by overlapping miRNA modules converging on shared signaling pathways. As a result, inhibition or overexpression of individual miRNAs often triggers compensatory mechanisms that blunt therapeutic efficacy [27,28]. This network resilience may explain the limited durability observed in some preclinical miRNA interventions and emphasizes the need for systems-level approaches that account for miRNA target network dynamics rather than single-miRNA effects. Cell-type specificity remains a critical bottleneck. Most miRNAs are expressed across multiple immune and non-immune compartments, and their targets frequently differ depending on cellular context. Current delivery strategies still struggle to selectively modulate miRNA activity in defined immune subsets, such as tumor-associated macrophages or exhausted T cells, without affecting other immune populations [29,30]. This lack of precision raises concerns regarding immune-related adverse effects, particularly when miRNAs influence pathways involved in immune tolerance, inflammation, or autoimmunity.
The heterogeneity of extracellular vesicles (EVs) introduces additional complexity. EV populations differ widely in size, cellular origin, cargo composition, and biodistribution, yet are often collectively analyzed as a single entity. This heterogeneity complicates the interpretation of EV-associated miRNA function and limits reproducibility across studies [31,32]. Although the mechanisms governing selective miRNA loading into EVs are increasingly characterized, they are not yet fully understood and may vary substantially across tumor types and stress conditions [9]. Standardized EV isolation, characterization, and functional validation protocols remain urgently needed to translate EV-miRNA findings into clinical applications. From a translational perspective, safety and durability remain unresolved issues. Early clinical experiences with miRNA-based therapeutics have highlighted challenges related to off-target gene regulation, innate immune activation, and dose-limiting toxicities [33]. Unlike small molecules or monoclonal antibodies, miRNAs inherently regulate multiple targets, increasing the likelihood of pleiotropic effects. In the context of cancer immunotherapy, such broad activity may be advantageous in principle but poses substantial risks if not tightly controlled.
Finally, the field lacks robust predictive frameworks to identify which patients are most likely to benefit from miRNA-based immunomodulation. While circulating and EV-associated miRNAs show promise as biomarkers, their integration into clinical decision-making is still in its infancy [34]. Variability in sampling methods, normalization strategies, and temporal dynamics further complicates their clinical interpretation. These challenges reveal that the successful reprogramming of antitumor immunity through miRNAs will require a shift from reductionist strategies toward context-aware, cell-specific, and network-informed therapeutic designs. Addressing these conceptual gaps is essential for transforming miRNAs from powerful biological regulators into reliable clinical tools.
Translational and Clinical Perspectives
Despite compelling preclinical evidence supporting microRNA (miRNA)-mediated immune modulation, the clinical translation of miRNA-based strategies in cancer immunotherapy has progressed more slowly than anticipated. Notably, the miR-34a mimic MRX34 represented the first-in-human clinical attempt to restore tumor-suppressive miRNA activity systemically. In the phase I trial involving patients with advanced solid tumors, MRX34 demonstrated successful delivery to tumor tissues and dose-dependent modulation of target gene expression, providing early proof-of-concept for miRNA-based therapy. However, despite evidence of clinical activity, the trial was terminated prematurely due to serious immune-mediated adverse events, including severe inflammatory and cytokine-related toxicities [33,35]. These findings emphasize the limitations of applying conventional single-target therapeutic frameworks to miRNA-based interventions, particularly in the context of immune modulation.
Beyond MRX34, several miRNA-based therapeutics are advancing through early clinical and translational pipelines, highlighting both the potential and complexity of this therapeutic class [36]. The miR-16 mimic TargomiR has demonstrated feasibility and preliminary clinical activity in mesothelioma and other solid tumors, while cobomarsen, an inhibitor of miR-155, has been evaluated in hematological malignancies [37]. These efforts underscore growing interest in miRNA-targeted interventions but also reveal persistent challenges related to delivery efficiency, off-target effects, and durability of response. In addition, regulatory pathway considerations for oligonucleotide-based therapeutics, including safety monitoring, dose optimization, and long-term toxicity assessment, remain substantial [36]. Large-scale manufacturing, quality control, and cost-effective production of complex delivery platforms such as nanoparticles or extracellular vesicle–based carriers remain additional barriers to clinical translation [23,38]. Addressing these regulatory and scalability challenges will be essential for integrating miRNA therapeutics into mainstream oncology practice.
One of the most promising translational opportunities lies in combination strategies. Rather than functioning as standalone agents, miRNAs may be more effective when used to sensitize tumors or immune cells to existing immunotherapies, such as immune checkpoint inhibitors. Preclinical studies demonstrate that miRNA modulation can influence PD-L1 expression, T-cell exhaustion, and macrophage polarization, thereby enhancing responsiveness to checkpoint blockade [1,8]. Such combinatorial approaches may allow lower miRNA doses, reduce toxicity while preserving immunomodulatory efficacy.
Advances in delivery technologies are also reshaping the translational landscape. Engineered nanoparticles and bioengineered EVs have shown improved stability, biodistribution, and immune cell targeting compared with earlier platforms [14,16]. In particular, EV-based delivery systems offer intrinsic biocompatibility and the potential for cell-specific targeting, making them attractive candidates for immune-focused miRNA delivery. However, large-scale manufacturing of EV-based therapeutics remains substantially more complex than conventional small molecules or monoclonal antibodies. Challenges include low yield, batch-to-batch variability, cargo heterogeneity, purification standardization, and stringent regulatory requirements for clinical-grade production. Overcoming these scalability and quality-control limitations will be essential before EV-mediated miRNA delivery can achieve widespread clinical application. Beyond therapeutic applications, miRNAs are gaining traction as predictive and dynamic biomarkers for immunotherapy. Circulating and EV-associated miRNAs reflect real-time immune states and treatment-induced adaptations, providing advantages over static genomic biomarkers [34]. Integrating miRNA signatures with established biomarkers such as tumor mutational burden or PD-L1 expression may improve patient stratification and treatment monitoring, although standardized analytical pipelines are still needed. These translational findings suggest that the most immediate clinical impact of miRNAs may arise from adjunctive and biomarker-driven applications, rather than monotherapy. Strategic integration with existing immunotherapeutic frameworks, coupled with continued refinement of delivery platforms, will be essential for realizing the clinical potential of miRNA-based immune reprogramming.
Despite their promise as dynamic biomarkers, several technical and translational challenges must be addressed before routine clinical implementation. Variability in sample processing and normalization strategies remains a major limitation, as circulating miRNA measurements can be influenced by hemolysis, RNA extraction efficiency, and differences in reference controls. In addition, inter-platform variability across sequencing, qPCR, and microarray-based detection systems complicate cross-study comparisons and standardization. For extracellular vesicle–associated miRNAs, inconsistencies in EV isolation, characterization, and cargo quantification further limit reproducibility. Establishing robust clinical validation frameworks, including standardized protocols, longitudinal sampling strategies, and multi-cohort validation, will be essential for integrating miRNA-based biomarkers into clinical decision-making for cancer immunotherapy.
Future Directions and Emerging Concepts
The next phase of microRNA (miRNA)-based immune reprogramming will require a decisive shift from single-miRNA interventions toward context-aware and cell-specific strategies. Increasing evidence indicates that therapeutic efficacy will depend less on the identity of individual miRNAs and more on their spatiotemporal regulation within defined immune compartments. Advances in single-cell and spatial technologies are expected to play a central role in this transition by enabling precise mapping of miRNA activity across immune cell states, thereby guiding rational therapeutic design.
One promising direction is the development of cell-type restricted miRNA delivery platforms. Engineered EVs, ligand-decorated nanoparticles, and immune cell–derived vesicles offer the potential to selectively target macrophages, dendritic cells, or exhausted T cells while minimizing systemic exposure. Such approaches may allow fine-tuning of immune responses rather than global immune activation or suppression, addressing a key limitation of earlier miRNA therapies. Emerging synthetic biology and genome-editing tools are also poised to reshape the field. CRISPR-based modulation of miRNA loci or miRNA–target interactions could enable durable and programmable immune reprogramming, particularly when combined with inducible or reversible control systems. These strategies may be especially valuable for dissecting complex miRNA networks and validating therapeutic targets prior to clinical translation.
From a translational standpoint, future miRNA-based interventions are likely to be most effective as part of multimodal immunotherapeutic regimens. Integrating miRNA modulation with immune checkpoint blockade, adoptive cell therapies, or cancer vaccines may help overcome resistance mechanisms and enhance therapeutic durability. In parallel, the incorporation of circulating and EV-associated miRNAs into biomarker-driven treatment algorithms could facilitate patient stratification and real-time monitoring of immune responses. All these emerging concepts point toward a future in which miRNAs function not as isolated therapeutic agents but as precision regulators embedded within broader immunotherapeutic frameworks. Realizing this vision will require continued integration of systems biology, delivery engineering, and clinical insight to translate miRNA biology into actionable immunotherapy strategies.
Concluding Perspective
MicroRNAs have emerged as powerful regulators of antitumor immunity, capable of shaping immune checkpoint signaling, immune cell function, and intercellular communication within the tumor microenvironment. As highlighted throughout this commentary, recent advances have deepened our understanding of the complexity and context dependence of miRNA-mediated immune regulation, while also exposing critical conceptual and translational challenges that must be addressed to fully realize their therapeutic potential. Moving forward, the successful application of miRNA-based strategies in cancer immunotherapy will depend on embracing precision, integration, and context awareness. Rather than targeting individual miRNAs in isolation, future approaches must account for network-level regulation, immune cell specificity, and dynamic tumor–immune interactions. The integration of miRNA modulation with established immunotherapeutic modalities, coupled with advances in delivery technologies and biomarker-driven patient stratification, offers a promising path toward durable and safe immune reprogramming. In this evolving landscape, miRNAs are best viewed not as standalone therapeutics but as versatile molecular tools that can be strategically deployed to fine-tune antitumor immune responses. Continued interdisciplinary efforts bridging molecular biology, immunology, and clinical oncology will be essential for translating these insights into effective and personalized cancer immunotherapies.
Funding
This work received no external funding.
Conflicts of Interest
The authors declare no conflicts of interest.
References
2. Mangang NL, Gargasz SK, Murugan SG, Kumar M, Shukla GC, Vijayaraghavalu S. MicroRNAs Modulating Cancer Immunotherapy Mechanisms and Therapeutic Synergies. Cancers (Basel). 2025 Dec 13;17(24):3978.
3. Kuang L, Wu L, Li Y. Extracellular vesicles in tumor immunity: mechanisms and novel insights. Mol Cancer. 2025 Feb 14;24(1):45.
4. Kim HI, Park J, Zhu Y, Wang X, Han Y, Zhang D. Recent advances in extracellular vesicles for therapeutic cargo delivery. Exp Mol Med. 2024 Apr;56(4):836–49.
5. Lin X, Yue L, Cheng K, Rao L. Engineering cellular vesicles for immunotherapy. Acc Mater Res. 2025 Jan 22;6(3):327–39.
6. Huemer F, Leisch M, Geisberger R, Zaborsky N, Greil R. miRNA-Based Therapeutics in the Era of Immune-Checkpoint Inhibitors. Pharmaceuticals (Basel). 2021 Jan 26;14(2):89.
7. Shi W, Zhang Z, Xu X, Tian Y, Feng L, Huang X, et al. Single-cell and spatial transcriptomics integration: new frontiers in tumor microenvironment and cellular communication. Front Immunol. 2025 Oct 2;16:1649468.
8. Di Martino MT, Riillo C, Scionti F, Grillone K, Polerà N, Caracciolo D, et al. miRNAs and lncRNAs as Novel Therapeutic Targets to Improve Cancer Immunotherapy. Cancers (Basel). 2021 Mar 30;13(7):1587.
9. Villarroya-Beltri C, Baixauli F, Gutiérrez-Vázquez C, Sánchez-Madrid F, Mittelbrunn M. Sorting it out: regulation of exosome loading. Semin Cancer Biol. 2014 Oct;28:3–13.
10. Shurtleff MJ, Temoche-Diaz MM, Karfilis KV, Ri S, Schekman R. Y-box protein 1 is required to sort microRNAs into exosomes in cells and in a cell-free reaction. Elife. 2016 Aug 25;5:e19276.
11. Zhang L, Zhang S, Yao J, Lowery FJ, Zhang Q, Huang WC, et al. Microenvironment-induced PTEN loss by exosomal microRNA primes brain metastasis outgrowth. Nature. 2015 Nov 5;527(7576):100–4.
12. Challagundla KB, Wise PM, Neviani P, Chava H, Murtadha M, Xu T, et al. Exosome-mediated transfer of microRNAs within the tumor microenvironment and neuroblastoma resistance to chemotherapy. J Natl Cancer Inst. 2015 May 13;107(7):djv135.
13. Whiteside TL. Exosomes and tumor-mediated immune suppression. J Clin Invest. 2016 Apr 1;126(4):1216–23.
14. Zhou Q, Xiang J, Qiu N, Wang Y, Piao Y, Shao S, et al. Tumor Abnormality-Oriented Nanomedicine Design. Chem Rev. 2023 Sep 27;123(18):10920–89.
15. O'Brien K, Breyne K, Ughetto S, Laurent LC, Breakefield XO. RNA delivery by extracellular vesicles in mammalian cells and its applications. Nat Rev Mol Cell Biol. 2020 Oct;21(10):585–606.
16. Kamerkar S, LeBleu VS, Sugimoto H, Yang S, Ruivo CF, Melo SA, et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature. 2017 Jun 22;546(7659):498–503.
17. Zhang Q, Pan J, Xiong D, Zheng J, McPherson KN, Lee S, et al. Aerosolized miR-138-5p and miR-200c targets PD-L1 for lung cancer prevention. Front Immunol. 2023 Jul 13;14:1166951.
18. Yadav R, Khatkar R, Yap KC, Kang CY, Lyu J, Singh RK, et al. The miRNA and PD-1/PD-L1 signaling axis: an arsenal of immunotherapeutic targets against lung cancer. Cell Death Discov. 2024 Sep 29;10(1):414.
19. Markopoulos GS, Roupakia E, Tokamani M, Alabasi G, Sandaltzopoulos R, Marcu KB, et al. Roles of NF-κB Signaling in the Regulation of miRNAs Impacting on Inflammation in Cancer. Biomedicines. 2018 Mar 30;6(2):40.
20. Haghikia A, Hoch M, Stapel B, Hilfiker-Kleiner D. STAT3 regulation of and by microRNAs in development and disease. JAKSTAT. 2012 Jul 1;1(3):143–50.
21. Longo F, Gattuso G, Spoto G, Ricci D, Vitale ACV, Lavoro A, et al. The multifaceted role of microRNAs in colorectal cancer: pathogenesis and therapeutic implications. Noncoding RNA Res. 2025 May 23;14:65–95.
22. Zuo H, Liao A, Shen S, Wang Y, Yang X, Weng S, et al. microRNA-34 mediates a negative feedback loop in the JAK-STAT pathway to attenuate immune overactivation in an invertebrate. Cell Commun Signal. 2025 May 17;23(1):228.
23. Salunkhe SS, Ghatage T, Prabhakar BS. Multi-oncogene targeting in cancer therapy: A miRNA-driven pharmacological approach. Biochem Pharmacol. 2025 Dec;242(Pt 4):117417.
24. Vishnubalaji R, Shaath H, Al-Alwan M, Abdelalim EM, Alajez NM. Reciprocal interplays between MicroRNAs and pluripotency transcription factors in dictating stemness features in human cancers. Semin Cancer Biol. 2022 Dec;87:1–16.
25. O'Connell RM, Rao DS, Baltimore D. microRNA regulation of inflammatory responses. Annu Rev Immunol. 2012;30:295–312.
26. Mehta A, Baltimore D. MicroRNAs as regulatory elements in immune system logic. Nat Rev Immunol. 2016 Apr 28;16(5):279–94.
27. Inui M, Martello G, Piccolo S. MicroRNA control of signal transduction. Nat Rev Mol Cell Biol. 2010 Apr;11(4):252–63.
28. Vidigal JA, Ventura A. The biological functions of miRNAs: lessons from in vivo studies. Trends Cell Biol. 2015 Mar;25(3):137–47.
29. Riley RS, June CH, Langer R, Mitchell MJ. Delivery technologies for cancer immunotherapy. Nat Rev Drug Discov. 2019 Mar;18(3):175–96.
30. Roberts TC, Langer R, Wood MJA. Advances in oligonucleotide drug delivery. Nat Rev Drug Discov. 2020 Oct;19(10):673–94.
31. Théry C, Witwer KW, Aikawa E, Alcaraz MJ, Anderson JD, Andriantsitohaina R, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018 Nov 23;7(1):1535750.
32. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020 Feb 7;367(6478):eaau6977.
33. Beg MS, Brenner AJ, Sachdev J, Borad M, Kang YK, Stoudemire J, et al. Phase I study of MRX34, a liposomal miR-34a mimic, administered twice weekly in patients with advanced solid tumors. Invest New Drugs. 2017 Apr;35(2):180–8.
34. Havel JJ, Chowell D, Chan TA. The evolving landscape of biomarkers for checkpoint inhibitor immunotherapy. Nat Rev Cancer. 2019 Mar;19(3):133–50.
35. Hong DS, Kang YK, Borad M, Sachdev J, Ejadi S, Lim HY, et al. Phase 1 study of MRX34, a liposomal miR-34a mimic, in patients with advanced solid tumours. Br J Cancer. 2020 May;122(11):1630–7.
36. Patutina O, Sen'kova A, Miroshnichenko S, Awad M, Markov O, Gladkikh D, et al. Targeted Inhibition of Oncogenic microRNAs miR-21, miR-17, and miR-155 Suppresses Tumor Growth and Modulates Immune Response in Colorectal Cancer. Pharmaceutics. 2026 Jan 18;18(1):122.
37. Dhuri K, Pradeep SP, Shi J, Anastasiadou E, Slack FJ, Gupta A, et al. Simultaneous Targeting of Multiple oncomiRs with Phosphorothioate or PNA-Based Anti-miRs in Lymphoma Cell Lines. Pharm Res. 2022 Nov;39(11):2709–20.
38. Wu HH, Leng S, Sergi C, Leng R. How MicroRNAs Command the Battle against Cancer. Int J Mol Sci. 2024 May 28;25(11):5865.