Commentary
This commentary will deal with the assessment of the aqueous permeability of biological membranes. Due to the hydrophobicity of the phospholipid bilayer, the membrane itself is not permeable to hydrophilic substances. However, the permeability of the natural biological membrane for small and electrically neutral water molecules is very high. The passive entry of water into cells is ensured by the presence of specific "water channels" in membranes, or aquaporins, the universal water channels, are responsible for rapid response of cell volume to changes in plasma tonicity. Aquaporins are independent protein complexes designed by nature for transmembrane transport of water along the osmotic gradient. They selectively allow water molecules to pass through membrane [1].
With their help water molecules enter and leave the cell, preventing the flow of ions and other soluble substances. Each cell has its own set of aquaporins, which is determined by the evolution of the organ and its function. Thus, water permeability depends directly on the number of aquaporins in the membrane [2]. For example, each erythrocyte contains up to 120-160 thousand molecules of aquaporin-1 (AQP1) [3]. Due to the high rate of water entry into the cell, its estimation is very difficult.
In 2014, the author published a method for assessing the aqueous permeability of erythrocyte membranes [4]. The method of gradient osmotic ektacytometry was used [5]. Gradient osmotic ektacytometry is a highly accurate and informative method recognized as the gold standard in the arsenal of diagnostic tools for studying the functional properties of erythrocytes. The deformability of the erythrocyte depends on the ratio of the surface and volume, or the shape of the cell, the internal viscosity and rigidity of the membrane. The classical profile of the osmotic deformability of native erythrocytes (osmoscan), which is a change in the deformability index (EI) of erythrocytes in the range of osmotic concentrations of the suspension medium, is usually characterized by the following parameters: integral deformability index (EImax) at isotonic osmolality O(EImax), EImin parameter at osmoscan inversion point Omin, Ohyper index, which varies linearly with MCHC, which serves as an estimate of the cytoplasmic viscosity, or the degree of hydration of hemoglobin in the erythrocyte and osmotic deformability range of erythrocytes (O=Ohyper-Omin).
The work evaluated the deformability index at the point of isotropic erythrocyte swelling Omin (Imin) with formation of additional water pores and blocking of existing pores was estimated. The essence of the method is that under hypoosmotic conditions water penetrates into erythrocytes along the concentration gradient. At the prehemolytic stage the erythrocyte has the shape of an isotropic sphere. The sphere has a maximum volume for a given surface area, a change in its morphology suggests two alternatives. If the volume is maintained, this transformation must be accompanied by an increase in surface area, and if the area is maintained, the volume must decrease. Thus, deformation of a spherical erythrocyte will lead to a decrease in its volume, since biological membranes are unstretchable [6,7]. Gradient ektacytometry shows that during the isotropic sphere stage the erythrocyte undergoes a transformation under the action of shear stress in Quette’s cell. An increase in hydrostatic pressure leads to a decrease in volume compared to the maximum critical one due to the exit of the liquid phase through the hydrophilic pores into the suspension medium. Thus, the index Imin clearly reflects the ability of the erythrocyte membrane to pass water molecules. Unfortunately, in the literature the parameter Imin of osmoscan is not discussed in detail.
To demonstrate the applicability of the proposed method for assessing the ability of the erythrocyte membrane to pass water molecules, we present experiments to study the dynamics of the erythrocyte deformation abilities formation in mature and immature newborn animals. The rheological properties of erythrocytes in Wistar rats and guinea pigs of both sexes were studied 1, 3, 10 days and 1, 2 and 3 months after birth. After decapitation of non-narcotized animals, blood was collected in tubes with EDTA. Rheological properties of erythrocytes were assessed by gradient ektacytometry at 37oC. Statistical processing of the results was performed using Microsoft Office Exel 2007. The data are presented as arithmetic mean values with their standard deviations. Significance of the changes found in the compared groups was determined using two-pair Student's test with unequal deviation.
Figures 1 and 2 show the averaged osmoscans of erythrocytes in postnatal ontogeny of the studied animal species. Digital data are presented in Table 1.
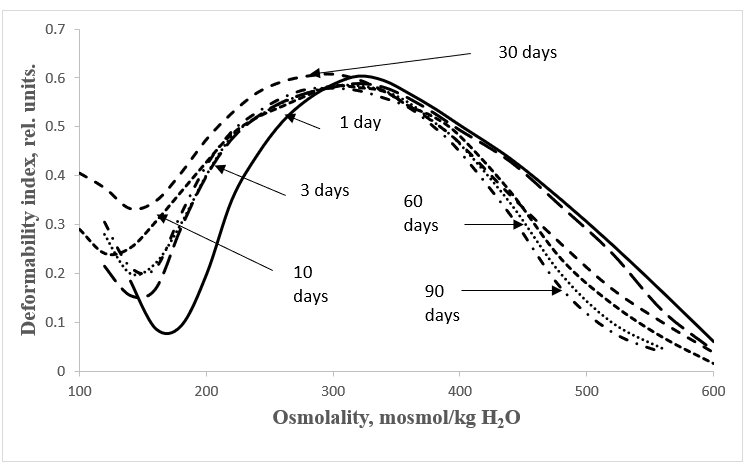
Figure 1: Erythrocyte osmoscans of Wistar rats at different stages of postnatal development.
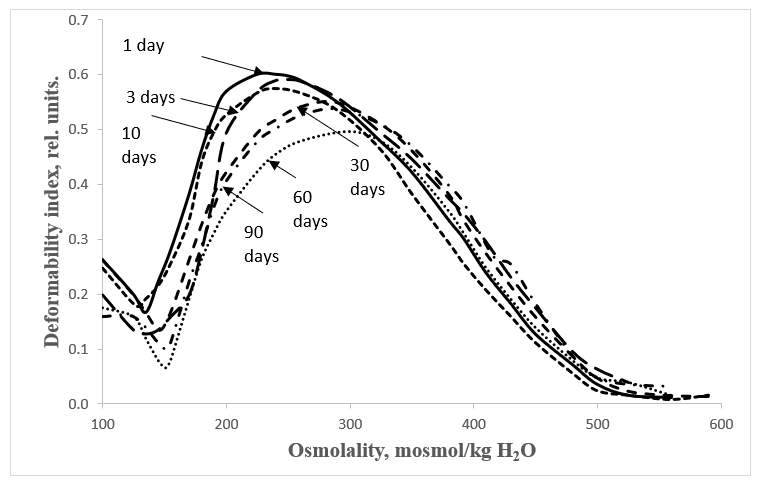
Figure 2: Erythrocyte osmoscans of guinea pigs at different stages of postnatal development.
| Age, days | Wistar rats | guinea pigs |
|---|---|---|
| 1 | 0,076 ± 0,004, n=5 | 0,207 ± 0,022, n=7 |
| 3 | 0,150 ± 0,010 ¥, n=5 | 0,128 ± 0,024 *, n=6 |
| 10 | 0,238 ± 0,015¥, n=5 | 0,179 ± 0,026, n=11 |
| 30 | 0,330 ± 0,018¥, n=5 | 0,135 ± 0,041, n=8 |
| 60 | 0,196 ± 0,016¥, n=5 | 0,066 ± 0,020¥, n=6 |
| 90 | 0,200 ± 0,020¥, n=5 | 0,100 ± 0,007 ¥, n=4 |
Note: n - number of animals; * - p<0.05; # - p<0.01; ¥ - p<0.001
Table 1: EImin parameter of Wistar rat and guinea pig erythrocytes at different stages of postnatal ontogenesis (M ± σ).
The presented data show that water permeability of erythrocyte membranes during postnatal ontogenesis in rats as mature newborn animals has a reliable tendency to increase, and in immature newborn pigs - to decrease. This pattern is demonstrated more clearly in the histograms presented in Figure 3.
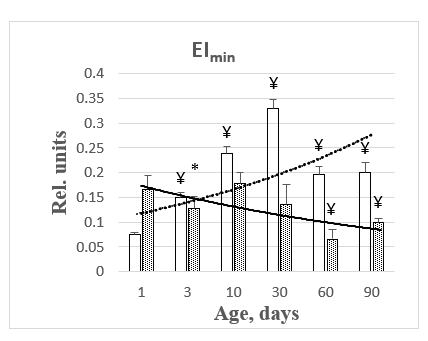
Figure 3: Histograms of changes in EImin parameter during maturation of rats and pigs.
Discussion
Osmoscan is a reflection of the superposition contribution of deformation changes of all the form elements of blood samples under study in a wide range of osmolality of the suspension medium. Analysis of the literature confirms the increase in aquaporin expression in cell membranes during mammalian ontogenesis. Thus, the level of AQP1 increased fivefold from the last gestational day to the first postnatal day and persisted at high levels into adulthood [8,9]. The content of AQP1 detected in the rat myometrium increases at the beginning of early pregnancy from day 1 to day 6 from the moment of conception [10]. The level of AQP1 in erythrocytes increases even after birth [11,12]. Compared to adults, it has been shown that human fetuses in the second and third trimesters contain less AQP1 and lower osmotic permeability to water [13]. Our message shows that the expression of the aquaporin pool in the membranes of rat erythrocytes, as immature animals, increases in postnatal ontogenesis. Unfortunately, systematic data on blood rheology in the ontogenesis of mature animals such as guinea pigs, starting from birth, are not available in the available literature at all. This report shows that guinea pig erythrocytes already possess a fully formed pool of AQP1 aquaporins by birth, which is gradually lost during the process of maturation. In this connection, it should be mentioned that the deformational properties of blood erythrocytes in both species of the studied animals also decrease during their maturation and, consequently, aging. This is evidenced by the changes in the integral deformability index (EImax).
Currently, a promising direction in molecular biology is the search for aquaporin inhibitors and activators [14]. Artificial modulators of aquaporins will find wide application in prevention in preventing heart diseases [15-17], treatment of blood diseases [18], and tumor processes [19-21]. Thus, in the context of ongoing research, the proposed method for assessing the water permeability of erythrocyte membranes seems very promising.
References
2. Zeidel ML, Nielsen S, Smith BL, Ambudkar SV, Maunsbach AB, Agre P. Ultrastructure, pharmacologic inhibition, and transport selectivity of aquaporin CHIP in proteoliposomes. Biochemistry. 1994 Feb 1;33(6):1606-15.
3. Denker BM, Smith BL, Kuhajda FP, Agre P. Identification, purification, and partial characterization of a novel Mr 28,000 integral membrane protein from erythrocytes and renal tubules. Journal of Biological Chemistry. 1988 Oct 25;263(30):15634-42.
4. Katyukhin LN. A method for evaluation of membrane permeability for water by the erythrocyte osmotic deformability profiles. Bulletin of Experimental Biology and Medicine. 2014 May 1;157(1):116-9.
5. Johnson RM. [2] Ektacytometry of red blood cells. InMethods in Enzymology 1989 Jan 1 (Vol. 173, pp. 35-54). Academic Press.
6. Ivens I, Skaylack R. Mechanics and thermodynamics of biological membranes. M.: Mir, 1982.–304 s.[In Russian]. 1982.
7. Hennis R. Biomembranes. Molecular Structure and Functions. 1997. M. Mir: 624 p.
8. Tsunoda SP, Wiesner B, Lorenz D, Rosenthal W, Pohl P. Aquaporin-1, nothing but a water channel. Journal of Biological Chemistry. 2004 Mar 19;279(12):11364-7.
9. King LS, Nielsen S, Agre P. Aquaporin-1 water channel protein in lung: ontogeny, steroid-induced expression, and distribution in rat. The Journal of Clinical Investigation. 1996 May 15;97(10):2183-91.
10. Lindsay LA, Murphy CR. Aquaporin-1 increases in the rat myometrium during early pregnancy. Journal of Molecular Histology. 2004 Jan;35(1):75-9.
11. Natochin Yu.V, Tkachuk VA. Modern course of classical physiology. M: Geotar-Media, 2009. 384 p.
12. Orlov RS, Nozdrachev AD. Normal Physiology: Textbook. M: Geotar-Media. 2005. 388 p.
13. Agre P, Smith BL, Baumgarten R, Preston GM, Pressman E, Wilson P, et al. Human red cell Aquaporin CHIP. II. Expression during normal fetal development and in a novel form of congenital dyserythropoietic anemia. The Journal of Clinical Investigation. 1994 Sep 1;94(3):1050-8.
14. Benga G. The first discovered water channel protein, later called aquaporin 1: molecular characteristics, functions and medical implications. Molecular Aspects of Medicine. 2012 Oct 1;33(5-6):518-34.
15. Song D, Yang Y, He N, Tian X, Sang DS, Li YJ. The involvement of AQP1 in myocardial edema induced by pressure overload in mice. Eur Rev Med Pharmacol Sci. 2018 Aug 1;22(15):4969-74.
16. Ding FB, Yan YM, Huang JB, Mei J, Zhu JQ, Liu H. The involvement of AQP1 in heart oedema induced by global myocardial ischemia. Cell Biochemistry and Function. 2013 Jan;31(1):60-4.
17. Li L, Weng Z, Yao C, Song Y, Ma T. Aquaporin-1 deficiency protects against myocardial infarction by reducing both edema and apoptosis in mice. Scientific Reports. 2015 Sep 8;5(1):13807.
18. Chow P, Cox C, Pei J, Anabaraonye N, Nourmohammadi S, Henderson S, et al. Inhibition of the Aquaporin-1 Cation Conductance by Selected Furan Compounds Reduces Red Blood Cell Sickling. Frontiers in Pharmacology. 2021 Jan 1;12:794791.
19. Wang Y, Fan Y, Zheng C, Zhang X. Knockdown of AQP1 inhibits growth and invasion of human ovarian cancer cells Retraction in/10.3892/mmr. 2021.12503. Molecular Medicine Reports. 2017 Oct 1;16(4):5499-504.
20. Simone L, Gargano CD, Pisani F, Cibelli A, Mola MG, Frigeri A, et al. Aquaporin-1 inhibition reduces metastatic formation in a mouse model of melanoma. Journal of Cellular and Molecular Medicine. 2018 Feb;22(2):904-12.
21. Titovets EP. Human and animal aquaporins: fundamental and clinical aspects. Minsk: Belorus. Nauka, 2007. 239 p.