Abstract
Human dopamine transporter gene (SLC6A3) is one of the neurotransmission genes that plays an important role in controlling the behavior and psychological reactions toward various physical activities. The aim of the present study is to evaluate the associations between SLC6A3 3’-UTR VNTR (3’VNTR) genotypes and eight personality traits in Egyptian athletes. One hundred athletes (50 males and 50 females) and one hundred non-athletes participated in this investigation either in individual or team games. The Freiburg Personality inventory-Revised questionnaire was applied to athletes to evaluate personality traits according to the scale, at a scheduled time, using descriptive analysis. Genotyping for SLC6A3 3’-UTR VNTR polymorphism in athletes was realized to detect the different genotypes by PCR amplifications using desired primers. Three different genotypes of the 3’VNTR polymorphism; namely (9/9), (9/10), (10/10) were identified. The 9/10 and 10/10 genotypes of SLC6A3 gene were significantly higher in players than those in the control group. There were no statistically significant differences in personality traits values among males and females. The Cronbach’s alpha stability coefficient for four items is higher than 0.7 suggesting that these items (Neuroticism, Aggressiveness, Depression and Excitability) have a good reliability and support the used personality dimensions for this investigation. Our findings revealed a significant association between the genotypes of 3’VNTR and the personality traits in Egyptian athletes. These data shed light on the role of SLC6A3 gene in athletes’ behavioral performance and could serve as the basis for further characterization in the large number of Egyptian athletes.
Keywords
SLC6A3 gene, 40 bp-VNTR polymorphism, DAT1, Personality traits, Athletes, Egyptian population
Introduction
Personality traits are the result of hereditary, human health and disease and environmental influence [1]. The natural genetic variants in the human genome displayed an effective implication in the individual behavior that could contribute to the level of athletic performance and related physical activity. The importance of personality traits evaluation and their interactions with athletes’ performance and expecting behavior were emphasized [2-4]. Twin and adoption studies have revealed that the heritability influences on human personality are ranged from 30 to 60%. Besides, many of the genetic variants have a possible impact on the development, but few have been identified [5]. Technical advances in DNA genotyping and sequencing have enabled the detection of the natural genetic variants that may have an effective association or impact on the level of athletic performance [6].
Numerous candidate genes with putative roles in sport performance have been suggested [7]. The human dopamine transporter gene plays an important role in controlling the behavior and psychological reactions toward various physical activities [8]. Dopamine (DA) neurotransmission represents a main endophenotype for measuring the changes in executive processes between individuals which constitutes Self-regulation behavior [9]. Furthermore, it is also one of the essential monoamine neurotransmitters in the central nervous system to regulate motivation, emotion, and cognition [10].
The SLC6A3 gene coding for the dopamine transporter (DAT1) is located in chromosome 5p15.33 and consists of 15 exons and 14 introns. In the 15th exon, there is a variable number of tandem repeat (VNTR) of a 40-base pair (bp) in the 3′-untranslated region (UTR) ranging from 3 to 13 times [11]. In fact, the 9- and 10-repeat polymorphisms are the most frequent which are suggested to be linked with DA-related psychiatric disorders and personality traits [12]. The SLC6A3 9-repeat allele is expressed lower than the 10-repeat allele in human cells [13-15], while another work stated the opposite [16]. However, the SLC6A3 gene expression in extra synaptic striatal fraction revealed that the 10-repeat allele is highly expressed, followed by a decrease in DA levels as confirmed with single-photon-emission computed tomography [13,17]. It is noteworthy that the importance of the SLC6A3 gene in regulating presynaptic DA homeostasis was characterized in transgenic mice lacking this gene [18]. Interestingly, mice without DAT1 present a spontaneous hyperlocomotion [19].
Allelic variants of the SLC6A3 gene have been implicated in human health and disease. For example, executive function components associated with SLC6A3 genotypes are observed in children with Attention-Deficit/Hyperactivity Disorder (ADHD) especially with the homozygous 10/10 genotype [20]. In contrast, Dresler et al. [21] found that the 9/9 & 9/10 repeat genotypes are associated with ADHD in patients with alterations in cognitive response and prefrontal brain function. Lee [2] explored the association between personality traits, environmental and physiological variables, and SLC6A3 genotypes in a group of 120 New Zealand students. This study showed that genetic effects for SLC6A3 on neurotic and sensation-seeking traits are masked by gender and ethnicity. In addition, one hypothesis is that DA transporters within the striatum in the brain moderated the dopaminergic system and psychological outcomes in children's self-regulatory capacities [9].
Several researchers have provided strong evidence for an association between the 40 bp-VNTR repeats of SLC6A3 gene and personality traits. Micha?owska-Sawczyn et al. [3] studied this association in Polish players and they proposed that differences in athletes’ personality traits are related to the allelic variants of the SLC6A3 gene. In this regard, the impact of new psychoactive substances (NPS) on personality traits, and its relationship with VNTR 3’-UTR polymorphisms were analyzed [4]. The 9/10 VNTR showed the highest score of neuroticism in NPS users compared to control subjects. No previous studies have been conducted in Egypt to confirm and characterize this association. The aim of this investigation is therefore to assess the relationship between personality traits scales (using the Freiburg Personality inventory- Revised (FPI-R) Questionnaire) with allele and genotype frequencies of the 40 base pair SLC6A3 VNTR 3’ UTR polymorphism in Egyptian athletes versus control.
Material and Methods
Subjects
One hundred athletes (50 females and 50 females) and one hundred healthy controls (non-athletes) participated in this investigation either in individual or team games. Participants were aged from 18 to 21 years old. The athletes blood samples were chosen according to the following criteria: (a) first-class players and Egyptian team; (b) players to be registered in appropriate sport federation; (c) players should be trained continuously for two years at least; (d) have participated in tournaments organized by sport unions. The Freiburg Personality inventory Revised (FPI-R) questionnaire was administered to athletes to evaluate personality traits scales on a typical day of training using the descriptive approach due to its relevance to the nature of the study [22]. Furthermore, a control group was defined as 50 men and 50 women to realize comparison with athletes in SLC6A3 alleles and genotypes frequencies.
Blood sampling and DNA Isolation
Approximately 5 mL of blood was collected from the peripheral blood of each player into EDTA tubes. Genomic DNA was isolated from blood using the QIAamp DNA extraction kit (Qiagen, Germany) according to the manufacturer's protocol. DNA was eluted in deionized water and then stored at -20°C until genotyping analyses. The quality of DNA was determined in a Nanodrop spectrophotometer (ThermoScienti?c, USA) at 260 nm and 280 nm absorbance ratios. A ratio of 1.8-2.0 indicates purity of DNA samples. Moreover, DNA samples were electrophoresed on a 1.5% agarose gel to confirm the integrity and quality of the extracted DNA. A ladder 1 kb DNA marker (ThermoScienti?c, USA) was used as a molecular weight reference.
Genotyping for 40 bp-VNTR polymorphism of SLC6A3 gene
The 40 base pair SLC6A3 3’-UTR VNTR genotypes were detected by PCR amplification using targeted primers: F- 5’TGTGGTGTAGGGAACGGCCTGAG-3’ (forward) and R- 5’CTTCCTGGAGGTCACGGCTCAAGG-3’ (reverse) [9]. The mixture of PCR reaction was applied in a total volume of 25 μl containing 100 ng DNA, 20 pmol of each primer, 200 mmol/L dNTPs, 1x reaction buffer, 1.5 mmol/L MgCl2 and 1U of Taq DNA polymerase (Promega, USA). The amplification was realized on the following conditions: an initial denaturation of 5 min at 95°C, followed by 50 cycles of denaturation (95°C, 30s), annealing (63°C, 30 s), and elongation (68°C, 45 s), and a final elongation step at 72°C for 5 min. The PCR products were separated by electrophoresis on a 2.5 % agarose gel.
The Freiburg Personality inventory- Revised (FPI-R) scales analysis
FPI-R scales were applied to detect personality profiles of studied athletes [23] and were translated and validated for the Egyptian population in Arabic by Allawi et al. [22]. These personality parameters consist of eight traits namely; neuroticism, aggressiveness, depression, excitability, social orientation, security, self-direction, and emotionality. The FPI is composed of 56 questions classified by eight personality traits and each scale, consisting of seven statements. Moreover, participants were asked to choose an answer for each statement according to their opinions, on a two-point Likert scale ranging from totally agree to nothing agree. The positive answer takes (2) points with totally agree and (1) point with disagree whereas the negative answer takes (1) point with totally agree and (2) points with disagree respectively.
Statistical analyses
The data were analyzed using the SPSS software (Ver. 18.0 IBM). The normality of data distribution was tested with the Shapiro-Wilk test. To assess differences in personality variables between studied players and control subjects, t-tests were performed. A Spearman correlation coefficient and Cronbach’s alpha stability coefficient were used to measure reliability, or internal consistency. SLC6A3 genotype and allele frequency comparison between groups were performed using a chi-square test. Significance level was considered with a p-value of less than 0.05. Hardy–Weinberg equilibrium (HWE) was performed in order to determine genotypes frequency and their distribution in the studied subjects.
Results
Slc6A3 3'-UTR VNTR genotypes
Two main alleles of SLC6A3 3’-UTR VNTR genotypes were presented in athletes under study. These alleles were: the 9-repeat allele with a length of 440 bp and the 10-repeat allele with a molecular length of 480 bp are shown in Figure 1. The distribution of the SLC6A3 3’-UTR VNTR genotype and allele frequencies were presented in Table 1. Three different genotypes were detected as follows: the homozygous 9/9 genotype (athletes 10.0% vs. controls 47.0%); the homozygous 10/10 (athletes 58.0% vs. controls 33.0%) and the heterozygous 9/10, (athletes 32.0% vs. controls 20.0%). The homozygous 10/10 genotype and heterozygous 9/10 of SLC6A3 gene frequencies were significantly higher in players than that in the control subjects (Table 1, p value =0.0001). Interestingly, genotype 9/9 was much higher in the control group than in the athletes group. The 9-allele frequency was approximately two-fold higher in the control group than in the athletes group (26.0%), whereas the frequency of the 10-allele may be higher in the athletes group (males and females, 74.0%) than in the control group (43.0%). All identified polymorphisms were in the Hardy–Weinberg equilibrium in the players’ samples but not in the control subjects as shown in Table 1.
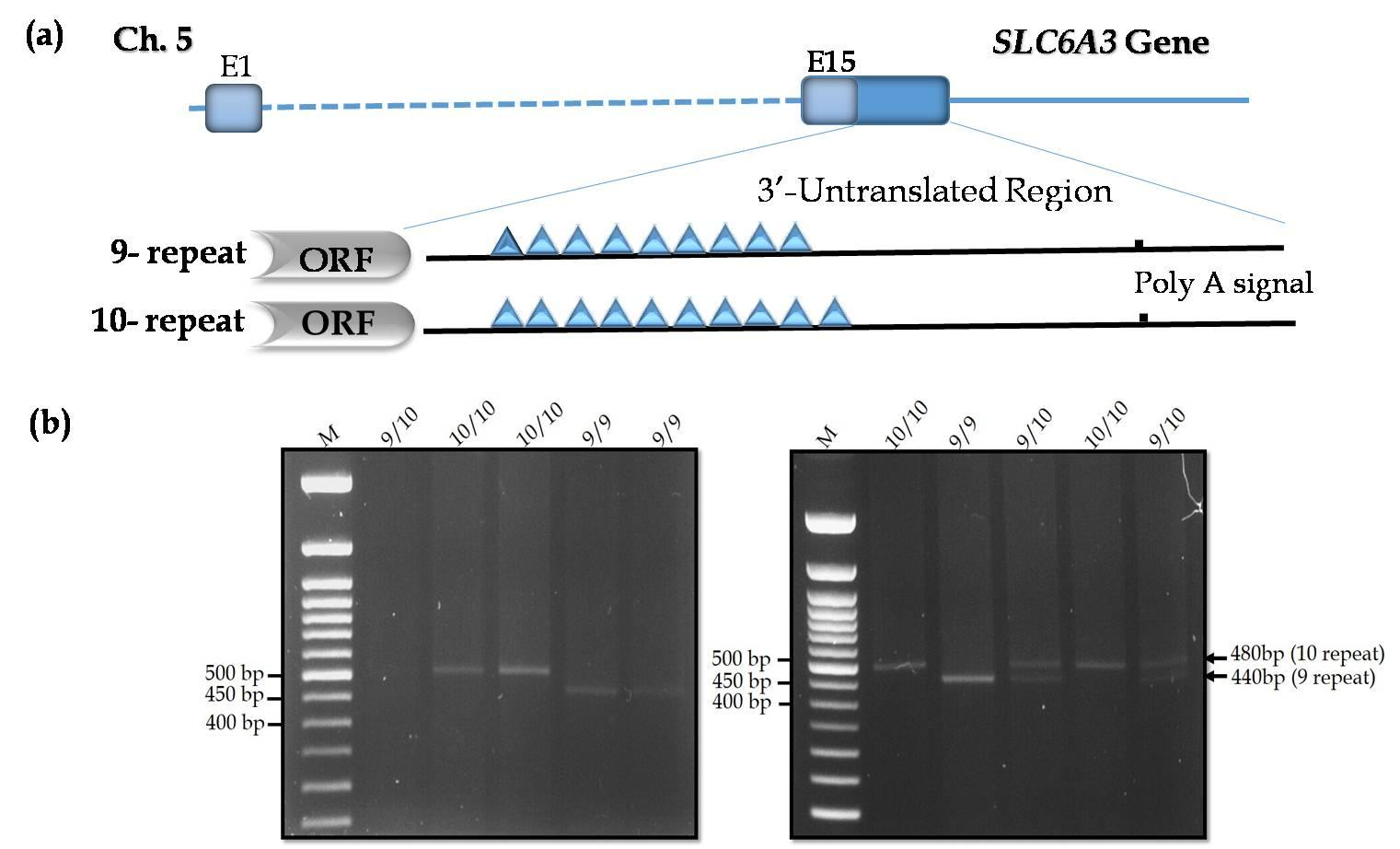
Figure 1. Allelic variants of SLC6A3, 3’-UTR VNTR polymorphism in the examined subjects. (a) schematic illustration of a partial SLC6A3 gene showing the position of VNTR variants identified in the 3’-UTR region with different allelic variants. Exons are presented by boxes (grey, coding sequences; bleu, untranslated sequences), ORF: Open Reading Frame; (b) first gel, Lanes 1: ladder (100bp), Lane 2: 9/10 repeat genotype; lane 3-4: 10/10 repeat genotype; lanes 5 & 6: 9/9 repeat genotype; second gel: Lanes 1: ladder (100bp), Lane 2 & 5: 10/10 repeat genotype; lane 3: 9/9 repeat genotype; lanes 4 & 6: 9/10 repeat genotype.
|
SLC6A3 3’-UTR VNTR |
||||||
|
Groups |
Genotypes |
Alleles |
||||
|
|
Total N |
9/9 N (%) |
9/10 N (%) |
10/10 N (%) |
9 N (%) |
10 N (%) |
|
Athletes |
100 |
10 (10%) |
32 (32%) |
58 (58%) |
52 (26%) |
148 (74%) |
|
Control |
100 |
47 (47%) |
20 (20%) |
33 (33%) |
114 (57%) |
86 (43%) |
|
Χ2 |
33.65 |
19.79 |
||||
|
p value |
0.0001 |
0.0001 |
||||
|
Χ2 (HWE)Athletes |
2.83 |
|||||
|
p value |
0.518 |
|||||
|
Χ2 (HWE)Control |
32.3 |
|||||
|
p value |
0.0000754 |
|||||
|
N: Number of participants, p: Statistical significance |
||||||
Personality traits scale
A trait is a dynamic, relatively stable general tendency to a certain type of behavior and its effect appears in a large number of life situations [24]. The FPI-R scales score was performed for athletes under evaluation and compared to their genotypes. Genetic association between SLC6A3 3’-UTR VNTR genotypes and FPI-R scales score is presented in Table 2. The 9/9 genotype for SLC6A3 gene was more prevalent with scores between 12 and 14 in Depression, Social orientation (80.00%) and Emotionality (80.00% each), followed by neuroticism (60.00%). The 10/10 genotype was more prevalent in Aggressiveness (59.60%), followed by Excitability and Self-direction (both 28.10%). Moreover, the heterozygote 9/10 genotype was more prevalent in Security (57.60%).
|
Scale no |
FPI-R scales |
Player Score SLC6A 3’-UTR VNTR Frequency and Percentages |
||||||||||||||
|
Score |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
|||||||||
|
Genotypes |
n |
% |
n |
% |
n |
% |
n |
% |
n |
% |
n |
% |
n |
% |
||
|
1 |
Neuroticism |
9/9 (n=10) |
|
|
|
|
2 |
20 |
2 |
20 |
4 |
40 |
|
|
2 |
20 |
|
9/10 (n=33) |
|
|
|
|
7 |
21.2 |
8 |
24.2 |
6 |
18.2 |
|
|
12 |
36.4 |
||
|
10/10 (n=57) |
|
|
|
|
18 |
31.6 |
16 |
28.1 |
16 |
28.1 |
|
|
7 |
12.3 |
||
|
2 |
Aggressiveness |
9/9 (n=10) |
|
|
2 |
20 |
|
|
4 |
40 |
|
|
4 |
40 |
|
|
|
9/10 (n=33) |
|
|
12 |
36.4 |
|
|
6 |
18.2 |
|
|
15 |
45.5 |
|
|
||
|
10/10 (n=57) |
|
|
7 |
12.3 |
|
|
16 |
28.1 |
|
|
34 |
59.6 |
|
|
||
|
3 |
Depression |
9/9 (n=10) |
|
|
|
|
|
|
2 |
20 |
4 |
40 |
|
|
4 |
40 |
|
9/10 (n=33) |
|
|
|
|
|
|
8 |
24.2 |
6 |
18.2 |
|
|
19 |
57.6 |
||
|
10/10 (n=57) |
|
|
|
|
|
|
16 |
28.1 |
16 |
28.1 |
|
|
25 |
43.9 |
||
|
4 |
Excitability |
9/9 (n=10) |
4 |
40 |
|
|
4 |
40 |
|
|
2 |
20 |
|
|
|
|
|
9/10 (n=33) |
6 |
18.2 |
|
|
19 |
57.6 |
|
|
8 |
24.2 |
|
|
|
|
||
|
10/10 (n=57) |
16 |
28.1 |
|
|
25 |
43.9 |
|
|
16 |
28.1 |
|
|
|
|
||
|
5 |
Social orientation |
9/9 (n=10) |
|
|
|
|
|
|
2 |
20 |
6 |
60 |
|
|
2 |
20 |
|
9/10 (n=33) |
|
|
|
|
|
|
8 |
24.2 |
18 |
54.5 |
|
|
7 |
21.1 |
||
|
10/10 (n=57) |
|
|
|
|
|
|
16 |
28.1 |
23 |
40.4 |
|
|
18 |
31.6 |
||
|
6 |
Security |
9/9 (n=10) |
|
|
|
|
6 |
60 |
|
|
4 |
40 |
|
|
|
|
|
9/10 (n=33) |
|
|
|
|
14 |
42.14 |
|
|
19 |
57.6 |
|
|
|
|
||
|
10/10 (n=57) |
|
|
|
|
32 |
56.1 |
|
|
25 |
43.9 |
|
|
|
|
||
|
7 |
Self-direction |
9/9 (n=10) |
|
|
2 |
20 |
6 |
60 |
|
|
2 |
20 |
|
|
|
|
|
9/10 (n=33) |
|
|
7 |
21.2 |
18 |
54.5 |
|
|
8 |
24.2 |
|
|
|
|
||
|
10/10 (n=57) |
|
|
18 |
31.6 |
23 |
40.4 |
|
|
16 |
28.1 |
|
|
|
|
||
|
8
|
Emotionality |
9/9 (n=10) |
2 |
20 |
|
|
|
|
|
|
4 |
40 |
|
|
4 |
40 |
|
9/10 (n=33) |
8 |
24.2 |
|
|
|
|
|
|
19 |
57.6 |
|
|
6 |
18.2 |
||
|
10/10 (n=57) |
16 |
28.1 |
|
|
|
|
|
|
25 |
43.9 |
|
|
16 |
28.1 |
||
There is no statistically significant relationship between the gene genotypes and the scales of personality traits, although it is close to the significant value in aggressiveness as shown in Table 3. The research sample is homogeneous on the Freiburg list of personality scales and that it has a similar level of these traits within each genotype (Table 4). The differences between males and females on the Freiburg personality list for the main research sample are not statistically significant (Table 5). Thus, the basic research sample will be treated as one group that includes males and females in the statistical analysis. The Cronbach's alpha stability coefficient for 4 items is higher than 0.7 suggesting that these items (Neuroticism, Aggressiveness, Depression and Excitability) have a good reliability and support the used personality dimensions for this investigation (Table 6).
|
FPI-R scales |
Spearman Correlation coefficient |
|
Neuroticism |
-0.195 |
|
Aggressiveness |
0.196 |
|
depression |
-0.074 |
|
Excitability |
0.023 |
|
Social orientation |
0.037 |
|
Security |
-0.070 |
|
Self-direction |
-0.037 |
|
Emotionality |
-0.023 |
|
*Significant at P<0.05 (tabular Spearman correlation coefficient = 0.197) |
|
|
Genotypes |
FPI-R scales |
Means ± SD |
Shapiro-Wilk Test |
|
|
Value |
indication |
|||
|
9/9 (n=10) |
Neuroticism |
11.8± 1.398 |
0.892 |
0.178 |
|
Aggressiveness |
11.4 ± 1.578 |
0.856 |
0.053 |
|
|
Depression |
12.6 ± 1.265 |
0.860 |
0.054 |
|
|
Excitability |
9.6 ± 1.578 |
0.870 |
0.059 |
|
|
Social orientation |
12.2 ± 1.033 |
0.887 |
0.176 |
|
|
Security |
10.8 ± 1.033 |
0.871 |
0.059 |
|
|
Self-direction |
10.8 ± 1.033 |
0.886 |
0.176 |
|
|
Emotionality |
12.00 ± 2.309 |
0.874 |
0.062 |
|
|
9/10 (n=33) |
Neuroticism |
12.06 ± 1.619 |
0.879 |
0.068 |
|
Aggressiveness |
11.18 ± 1.828 |
0.876 |
0.066 |
|
|
depression |
12.91 ± 1.331 |
0.896 |
0.186 |
|
|
Excitability |
10.12 ± 1.317 |
0.875 |
0.067 |
|
|
Social orientation |
12.18 ± 1.044 |
0.851 |
0.055 |
|
|
Security |
11.15 ± 1.004 |
0.890 |
0.184 |
|
|
Self-direction |
10.82 ± 1.044 |
0.851 |
0.055 |
|
|
Emotionality |
11.39 ± 2.091 |
0.857 |
0.057 |
|
|
10/10 (n=57) |
Neuroticism |
11.33 ± 1.272 |
0.880 |
0.187 |
|
Aggressiveness |
11.95 ± 1.42 |
0.888 |
0.195 |
|
|
depression |
12.6 ± 1.307 |
0.871 |
0.185 |
|
|
Excitability |
10.0 ±1.512 |
0.882 |
0.187 |
|
|
Social orientation |
12.35 ± 1.203 |
0.880 |
0.187 |
|
|
Security |
10.88 ± 1.001 |
0.890 |
0.195 |
|
|
Self-direction |
10.6 5± 1.203 |
0.891 |
0.195 |
|
|
Emotionality |
11.44 ± 2.322 |
0.885 |
0.191 |
|
|
FPI-R scales |
Male |
Female |
T- value |
|
Means ± SD |
Means ± SD |
||
|
Neuroticism |
11.62 ± 1.41 |
11.62 ± 1.47 |
0 |
|
Aggressiveness |
11.64 ± 1.59 |
11.64 ± 1.64 |
0 |
|
Depression |
12.66 ± 1.3 |
12.74 ± 1.32 |
-0.305 |
|
Excitability |
9.96 ± 1.48 |
10.04 ± 1.43 |
-0.275 |
|
Social orientation |
12.26 ± 1.12 |
12.3 ± 1.15 |
-0.176 |
|
Security |
10.92 ± 1.01 |
11 ± 1.01 |
-0.397 |
|
Self-direction |
10.74 ± 1.12 |
10.7 ± 1.15 |
0.176 |
|
Emotionality |
11.52 ± 2.27 |
11.44 ± 2.21 |
0.178 |
|
*Significant at P <0.05 (Tabular T = 1.984) |
|||
|
FPI-R sub-scales
|
Phrase (N) |
Cronbach's alpha stability coefficient |
Stability coefficient in case of item removed |
FPI-R sub-scales |
Phrase (N) |
Cronbach's alpha stability coefficient |
Stability coefficient in case of item removed |
|||
|
Sub-scale |
Total scale |
Sub-scale |
Total scale |
|
Sub-scale |
Sub-scale |
Total scale |
|||
|
Neuroticism |
3 |
0.784 |
0.778 |
0.676 |
0.488 |
Social orientation |
12 |
0.742 |
0.706 |
0.598 |
|
4 |
0.676 |
0.488 |
28 |
0.700 |
0.601 |
|||||
|
15 |
0.669 |
0.500 |
48 |
0.700 |
0.601 |
|||||
|
18 |
0.776 |
0.488 |
2 |
0.738 |
0.590 |
|||||
|
23 |
0.723 |
0.691 |
14 |
0.736 |
0.488 |
|||||
|
38 |
0.723 |
0.699 |
47 |
0.650 |
0.522 |
|||||
|
54 |
0.723 |
0.699 |
51 |
0.708 |
0.588 |
|||||
|
Aggressiveness |
7 |
0.779 |
0.778 |
0.710 |
0.738 |
Security |
1 |
0.738 |
0.636 |
0.587 |
|
10 |
0.778 |
0.706 |
20 |
0.682 |
0.598 |
|||||
|
26 |
0.702 |
0.753 |
29 |
0.708 |
0.488 |
|||||
|
27 |
0.702 |
0.570 |
42 |
0.708 |
0.488 |
|||||
|
41 |
0.702 |
0.570 |
43 |
0.640 |
0.600 |
|||||
|
44 |
0.697 |
0.731 |
45 |
0.675 |
0.689 |
|||||
|
49 |
0.703 |
0.728 |
56 |
0.654 |
0.679 |
|||||
|
Depression |
21 |
0.761 |
0.778 |
0.650 |
0.488 |
Self-direction |
9 |
0.725 |
0.648 |
0.621 |
|
25 |
0.650 |
0.488 |
11 |
0.701 |
0.618 |
|||||
|
34 |
0.650 |
0.488 |
16 |
0.713 |
0.732 |
|||||
|
37 |
0.760 |
0.602 |
22 |
0.715 |
0.567 |
|||||
|
40 |
0.760 |
0.602 |
24 |
0.705 |
0.770 |
|||||
|
52 |
0.760 |
0.575 |
30 |
0.665 |
0.699 |
|||||
|
55 |
0.759 |
0.583 |
50 |
0.676 |
0.699 |
|||||
|
Excitability |
5 |
0.800 |
0.778 |
0.710 |
0.748 |
Emotionality |
6 |
0.782 |
0.700 |
0.592 |
|
31 |
0.710 |
0.740 |
8 |
0.732 |
0.488 |
|||||
|
33 |
0.704 |
0.753 |
13 |
0.732 |
0.488 |
|||||
|
36 |
0.704 |
0.745 |
17 |
0.767 |
0.567 |
|||||
|
39 |
0.727 |
0.705 |
19 |
0.767 |
0.567 |
|||||
|
46 |
0.750 |
0.581 |
32 |
0.771 |
0.567 |
|||||
|
53 |
0.738 |
0.589 |
35 |
0.771 |
0.567 |
|||||
Discussion
Human athletic performance traits such as neuroticism, depression, social orientation, aggressiveness, excitability, self-direction, security, and emotionality are the result of the interaction between genetic and environmental influences, with polygenic basis in exercise-related traits [25]. This investigation highlights the role of 40 bp-VNTR polymorphisms of SLC6A3 gene in athlete’s personality traits and its influence on athletic performance. There are different factors affecting athletic performance that are well established: intensity and type of training, anthropometric characteristics as well as an important psychological component. However, the contribution of the genetic background has been less investigated [26]. Moreover, variation in personality parameters is one important component of human individuality and this variation is due to the interaction of interindividual differences in genes and environment [27].
The frequent alleles namely; the 9-repeat allele and the 10-repeat allele have effective effect on DAT1 expression due to DA reuptake from extracellular space [14,15,28-30]. Allelic variants could modify the gene expression profiling due to changes in regulatory sequences which affect the binding of transcription factors and related elements. Consequently, the gene product pattern will be modified [31]. In the present study, we identified that the 10-repeat allele was common in the athletes group and to be related to personality traits examined in the participants under study, whereas the 9-repeat allele was less common for the same group. It seems to be important for athletic performance. Bielinski et al. [32] showed an association between depression and angry-impulsive traits and the 9-repeat allele of SLC6A3 gene, which is in accordance with our results.
HWE tells geneticists where to look to find the causes of evolution in the population. Surprisingly, the population of Egyptian athletes is at the HWE suggesting an ideal population with assumptions of random mating, absence of selection or mutation and equal viability and fertility hold [33]. However, interestingly, the control population is not at HWE, suggesting a genetic drift or migration, which could alter genotype and allelic frequencies.
These findings showed that the effect of polymorphisms in the 40 bp-VNTR repeats of SLC6A3 gene may synergize and result in impaired dopaminergic signaling. The variation in athletes personality traits scale could be due to the difference in genotypes distribution of SLC6A3 gene and result in impaired DAT activity and DA signaling. Our results are in accordance with Miller & Madras [30] who provide experimental evidence that 40 bp-VNTR repeats of SLC6A3 gene may influence levels of DAT protein in the brain and affect human personality traits. Besides, dopamine plays an important role in several brain functions and is involved in the pathogenesis of several psychiatric and neurological disorders [34].
Athlete’s behavior tends to be stable to some extent and this is what is called “stability of behavior”, therefore the trait is a fixed style of behavior that distinguishes one individual from another [35]. Besides that, a new trend of personality investigation has shed the light on the significance of natural differences intraindividual which constitutes the essential base for various behavior and feeling across the changeable situation and time [36].
Genetic association between SLC6A3 3’-UTR VNTR genotypes and FPI-R scales score revealed that the 9/9 genotype was more common in neuroticism, depression and social orientation, whereas the 10/10 genotype was more common in aggressiveness, excitability and self-direction. Moreover, the heterozygote 9/10 genotype was more common in security and emotionality.
However, it can be concluded that studying athletic personality is important to identify dominant traits and how to deal with it, as this helps in: 1) giving the true indicators that it should be used in selecting juniors and players, which makes the sports training process more effective as it is focused on athletes who have the qualifications to succeed. 2) developing physical, skill and planning preparation programs aimed at gaining them motor abilities of the practiced sports activity. 3) determining the appropriate methods to follow in preparing athletes considering individual differences between the players. 4) studying the psychological aspect of an athlete's personality helps those in charge of his preparation in determining the size and intensity of the competitions that must be completed in the sports season.
The relationship between sport performance including creativity and neurotransmitter genes or brain-related genes such as the SLC6A3 3’-UTR VNTR polymorphism is not well characterized. The recent findings by Kitazawa et al. [37] present the influence of neurotransmitters in the CNS (central nerve system) and physical performance. Moreover, these results revealed that Dopamine could be responsible for the neurotransmission regulation in the CNS which will subsequently increase the athletic performance phenotypes. Such interested findings could prove to be valuable implications for future treatment strategies.
Recent investigations have emphasized the importance of personality traits evaluation and their relationship with athletes performances and expected behavior [2-4]. In this study, four personality traits seem to be important. Firstly, the aggressiveness trait is more prevalent in 10/10 genotype, followed by 9/10 genotype. This indicates that high score on this subscale refers to athletes who automatically submit physical, verbal, or imaginary aggression acts and respond emotionally and confront others with attacks and quarrels. They also characterized by impulsivity, inability to control themselves, lack of calmness and emotional immaturity. Moreover, three other traits are interesting: 1- the neuroticism trait is more prevalent in 9/9 genotype, followed by 9/10 genotype. This indicates that high score on this subscale distinguishes athletes who suffer from physical disorders (such as blood circulation, breathing, and digestion disorders, etc.), movement disorders (such as feeling pain), general psychosomatic disorders, sleep disturbances, fatigue, tension, rapid feeling of fatigue, sensitivity to weather changes, loss of comfort, and clarity of some nervous and physical manifestations associated with emotional arousal. 2- Depression trait is more prevalent in the 9/9 genotype, followed by 9/10 genotype. This indicates that a high score on this subscale distinguishes athletes who characterized by depression, mood swings, pessimism, feeling unhappy, dissatisfaction, fear, feeling unspecified fears, loneliness, lack of others understanding them, inability to focus, tendency to aggression against self, and feeling guilty. Low score on this subscale distinguish athletes who are of moderate temperament, the ability to focus, reassurance, self-confidence and contentment. 3- Excitability trait is more prevalent in 10/10 genotype, followed by 9/10 genotype. This indicates that the high score in this subscale distinguishes athletes who characterized by high arousal, high tension, inability to face normal daily frustrations, annoyance, impatience, anger, aggressive responses when frustrated, irritability, and of sensitivity.
Other studies showed a significant association between SLC6A3 40 bp-VNTR polymorphisms and suicidal behavior [38], ADHD disorder [39], and Eating Disorders with Binge Eating Behavior [40]. However, these data could provide a framework and new insights into the identification of additional candidate genes involved in the modulation of personality traits and sports performance, as well as the potential related disorders.
Conclusions
The current findings revealed a significant association between the genotypes of SLC6A3, 3’-UTR VNTR and personality traits in Egyptian athletes even with insufficient scales limitations. The 10 R-allele was more common in the athletes group than the 9 R-allele. This association will highlight the importance of SLC6A3 polymorphism as a brain-related gene in the variation of personality profile and its impact on the physical performances among athletes. Besides, this data could help in the classification of athletes and their behavior in order to find the effective genotype to improve athletic performances and personality parameters. Moreover, these data shed light on the role of SLC6A3 gene in athletes’ behavioral performance and could serve as the basis for further genetic screening and characterization in the large number of Egyptian athletes.
Ethics Statement
All subjects under the present study gave their written informed consent before obtaining any samples and performing any personality traits questionnaire. This study was reviewed and approved by the Department of Educational, Psychological and Social sciences, Faculty of Physical Education for Girls, Alexandria University.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding authors.
Authors Contributions
AE, contributed to full genetic analysis, and manuscript preparation; MB, collected athlete's samples, performed psychological Questionnaire of the athlete's personality, data analysis, and manuscript preparation. GP, supervising, and manuscript preparation; VL, is the corresponding author, supervising, and manuscript preparation. All authors read and approved the final version of this paper.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
2. Lee AS. Genes, beats and traits: A multi-marker exploration of personality. 2014. MSC Thesis, Victoria University of Wellington.
3. Michałowska-Sawczyn M, Lachowicz M, Grzywacz A, Suchanecka A, Chmielowiec J, Chmielowiec K, et al. Analysis of Polymorphic Variants of the Dopamine Transporter (DAT1) Gene Polymorphism and Personality Traits Among Athletes. Journal of Human Kinetics. 2020 Mar 1;72(1):79-89.
4. Chmielowiec J, Chmielowiec K, Masiak J, Pawłowski T, Larysz D, Grzywacz A. Analysis of Relationships between DAT1 Polymorphism Variants, Personality Dimensions, and Anxiety in New Psychoactive Substance (Designer Drug)(NPS) Users. Genes. 2021 Dec;12(12):1977.
5. Zwir I, Arnedo J, Del-Val C, Pulkki-Råback L, Konte B, Yang SS, et al. Uncovering the complex genetics of human character. Molecular Psychiatry. 2020 Oct;25(10):2295-312.
6. Lippi G, Longo UG, Maffulli N. Genetics and sports. British Medical Bulletin. 2010 Mar 1;93(1):27-47.
7. Raab M. Performance Psychology: A Guiding Framework for Sport Psychology. Handbook of Sport Psychology. 2020 Apr 14:1111-30.
8. Filonzi L, Franchini N, Vaghi M, Chiesa S, Marzano FN. The potential role of myostatin and neurotransmission genes in elite sport performances. Journal of Biosciences. 2015 Sep;40:531-7.
9. Cómbita LM, Voelker P, Abundis-Gutiérrez A, Pozuelos JP, Rueda MR. Influence of the SLC6A3-DAT1 gene on multifaceted measures of self-regulation in preschool children. Frontiers in Psychology. 2017 Jan 19;8:26.
10. Campbell NG, Shekar A, Aguilar JI, Peng D, Navratna V, Yang D, et al. Structural, functional, and behavioral insights of dopamine dysfunction revealed by a deletion in SLC6A3. Proceedings of the National Academy of Sciences. 2019 Feb 26;116(9):3853-62.
11. Vandenbergh DJ, Persico AM, Hawkins AL, Griffin CA, Li X, Jabs EW, et al. Human dopamine transporter gene (DAT1) maps to chromosome 5p15. 3 and displays a VNTR. Genomics. 1992 Dec 1;14(4):1104-6.
12. Kanno K, Ishiura S. Differential effects of the HESR/HEY transcription factor family on dopamine transporter reporter gene expression via variable number of tandem repeats. Journal of Neuroscience Research. 2011 Apr;89(4):562-75.
13. Heinz A, Goldman D, Jones DW, Palmour R, Hommer D, Gorey JG, et al. Genotype influences in vivo dopamine transporter availability in human striatum. Neuropsychopharmacology. 2000 Feb;22(2):133-9.
14. Mill J, Asherson P, Browes C, D'Souza U, Craig I. Expression of the dopamine transporter gene is regulated by the 3' UTR VNTR: Evidence from brain and lymphocytes using quantitative RT-PCR. American Journal of Medical Genetics. 2002 Dec 8;114(8):975-9.
15. VanNess SH, Owens MJ, Kilts CD. The variable number of tandem repeats element in DAT1 regulates in vitro dopamine transporter density. BMC Genetics. 2005 Dec;6:55.
16. van Dyck CH, Malison RT, Jacobsen LK, Seibyl JP, Staley JK, Laruelle M, et al. Increased dopamine transporter availability associated with the 9-repeat allele of the SLC6A3 gene. Journal of Nuclear Medicine. 2005 May 1;46(5):745-51.
17. Jacobsen LK, Staley JK, Zoghbi SS, Seibyl JP, Kosten TR, Innis RB, et al. Prediction of dopamine transporter binding availability by genotype: a preliminary report. American Journal of Psychiatry. 2000 Oct 1;157(10):1700-3.
18. Jones SR, Gainetdinov RR, Jaber M, Giros B, Wightman RM, Caron MG. Profound neuronal plasticity in response to inactivation of the dopamine transporter. Proceedings of National Academy of Sciences of the United States of America. 1998 Mar 31;95(7):4029-34.
19. Giros B. Jaber M, Jones SR, Wightman RM, Caron MG. Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature. 1996;379:606-12.
20. Karama S, Grizenko N, Sonuga-Barke E, Doyle A, Biederman J, Mbekou V, et al. Dopamine transporter 3'UTR VNTR genotype is a marker of performance on executive function tasks in children with ADHD. BMC Psychiatry. 2008 Dec;8:45.
21. Dresler T, Ehlis AC, Heinzel S, Renner TJ, Reif A, Baehne CG, et al. Dopamine transporter (SLC6A3) genotype impacts neurophysiological correlates of cognitive response control in an adult sample of patients with ADHD. Neuropsychopharmacology. 2010 Oct;35(11):2193-202.
22. Allawi MH. Encyclopedia of psychological tests for athletes. Cairo: Book publishing center. 1998; p 78-92.
23. Fahrenberg J, Hampel R, Selg H. Die revidierte form des freiburger persönlichkeitsinventars FPI-R. Diagnostica. 1985;31(1):1-21.
24. Zahran H. Mental health and psychotherapy. (4th edition). Cairo: Alam Al-Kutub. 2005; p.59.
25. Ginevičienė V, Utkus A, Pranckevičienė E, Semenova EA, Hall EC, Ahmetov II. Perspectives in sports genomics. Biomedicines. 2022 Jan 27;10(2):298-314.
26. Sanhueza JA, Zambrano T, Bahamondes-Avila C, Salazar LA. Association of anxiety-related polymorphisms with sports performance in Chilean long distance triathletes: a pilot study. Journal of Sports Science & Medicine. 2016 Dec;15(4):554-61.
27. Eaves L, Eysenck H. The nature of extraversion: a genetical analysis. Journal of Personality and Social Psychology. 1975 Jul;32(1):102-12.
28. Fuke S. Suo S, Takahashi N, Koike H, Sasagawa N, Ishiura S. The VNTR polymorphism of the human dopamine transporter (DAT1) gene affects gene expression. Pharmacogenomics J. 2001;1:152-6.
29. Michelhaugh SK, Fiskerstrand C, Lovejoy E, Bannon MJ, Quinn JP. The dopamine transporter gene (SLC6A3) variable number of tandem repeats domain enhances transcription in dopamine neurons. Journal of Neurochemistry. 2001 Dec;79(5):1033-8.
30. Miller GM, Madras BK. Polymorphisms in the 3'-untranslated region of human and monkey dopamine transporter genes affect reporter gene expression. Molecular psychiatry. 2002 Jan;7(1):44-55.
31. Müller F, Zaucker A, Tora L. Developmental regulation of transcription initiation: more than just changing the actors. Current Opinion in Genetics & Development. 2010 Oct 1;20(5):533-40.
32. Bieliński M, Jaracz M, Lesiewska N, Tomaszewska M, Sikora M, Junik R, et al. Association between COMT Val158Met and DAT1 polymorphisms and depressive symptoms in the obese population. Neuropsychiatric Disease and Treatment. 2017 Aug 18:2221-9.
33. Yu C, Zhang S, Zhou C, Sile S. A likelihood ratio test of population Hardy-Weinberg equilibrium for case-control studies. Genetic Epidemiology: The Official Publication of the International Genetic Epidemiology Society. 2009 Apr;33(3):275-80.
34. Ko JH, Strafella AP. Dopaminergic neurotransmission in the human brain: new lessons from perturbation and imaging. The Neuroscientist. 2012 Apr;18(2):149-68.
35. Darwish W. Sport Psychology: Theories and Applications. Alexandria: Sports World Foundation. Dar Al-Wafa’a for world of printing. 2014;p.156.
36. Jayawickreme E, Fleeson W, Beck ED, Baumert A, Adler JM. Personality dynamics. Personality Science. 2021 Aug 31;2:e6179.
37. Kitazawa H, Hasegawa K, Aruga D, Tanaka M. Potential genetic contributions of the central nervous system to a predisposition to elite athletic traits: state-of-the-art and future perspectives. Genes. 2021 Mar 5;12(3):371.
38. Rafikova E, Shadrina M, Slominsky P, Guekht A, Ryskov A, Shibalev D, et al. SLC6A3 (DAT1) as a novel candidate biomarker gene for suicidal behavior. Genes. 2021 Jun 4;12(6):861.
39. Grünblatt E, Werling AM, Roth A, Romanos M, Walitza S. Association study and a systematic meta-analysis of the VNTR polymorphism in the 3'-UTR of dopamine transporter gene and attention-deficit hyperactivity disorder. Journal of Neural Transmission. 2019 Apr 1;126:517-29.
40. Hirata T, Uemura T, Shinohara M, Hirano M. Association between Dopamine Transporter Gene (DAT1) Polymorphisms and Eating Disorders with Binge Eating Behavior. Open Journal of Psychiatry. 2017 Aug 16;7(4):329-43.